2.1. Численность популяции.
Популя́ция (от лат. populatio —
население) — это совокупность особей
одного вида, обитающих на определенной
территории, свободно скрещивающихся
между собой и частично или полностью
изолированных от других популяций.
Популяция
обладает только ей присущими особенностями:
численностью, плотностью, пространственным
распределением особей. Различают
возрастную, половую, размерную структуру
популяции. Соотношение разных по возрасту
и полу групп в популяции определяют ее
основные функции. Соотношение разных
возрастных групп зависит от двух причин:
от особенностей жизненного цикла вида
и от внешних условий.
Число
особей в популяции (численность популяции)
меняется со временем. Если условия
существования популяции благоприятны,
то рождаемость превышает смертность и
общее число особей в популяции растет
со временем. Назовем скоростью роста
популяции прирост числа особей в единицу
времени. Обозначим эту скорость v = v(t). В
“старых”, установившихся популяциях,
давно обитающих в данной местности,
скорость роста v (t) мала
и медленно стремится к нулю. Но если
популяция молода, ее взаимоотношения
с другими местными популяциями еще
не установились или существуют внешние
причины, изменяющие эти взаимоотношения,
например сознательное вмешательство
человека, то v (t) может
значительно колебаться, уменьшаясь
или увеличиваясь.[1]
Если известна скорость роста
популяции v t/),
то мы можем найти прирост численности
популяции за промежуток времени от tо
до Т. В
самом деле, из определения v(t) следует,
что эта функция является производной
от численности популяции N
(t) в
момент t, и,
следовательно, численность популяции
N (t) является первообразной
для v (t). Поэтому
N(t)
– N(t ![]() )
)
=  .
.
Известно,
что в условиях
неограниченных ресурсов
питания
скорость
роста многих популяций экспоненциальна,
т. е. v(t)
= ае ![]() . Популяция
. Популяция
в этом случае как бы “не стареет”. Такие
условия можно создать, например, для
микроорганизмов, пересаживая время от
времени развивающуюся культуру в новые
емкости с питательной средой. Применяя
формулу (1), в этом случае получим:
N(t)
= N(t ![]() )
)
+ a ![]() =
=
N(t ![]() )
)
+ ![]() e
e ![]()
![]() =
=
N(t ![]() )
)
+ ![]() (e
(e ![]() —
—
e ![]() )
)
По
формуле, подобной N(t) = N(t ![]() )
)
+ a ![]() =
=
N(t ![]() )
)
+ ![]() e
e ![]()
![]() =
=
N(t ![]() )
)
+ ![]() (e
(e ![]() —
—
e ![]() )
)
,
подсчитывают, в частности, численность
культивируемых плесневых грибков,
выделяющих пенициллин.
Пример задачи на численность популяции
Рассмотрим
микробиологическую задачу. Установим
закон изменения со временем (t) численности
бактерий (n),
помещенных в питательную среду.
Для составления дифференциального
уравнения, отражающего существование
бактерий в этих условиях, необходим
некоторый факт, который следует записать
в математической форме. На основании
экспериментальных данных и общих
соображений таким фактом может служить
утверждение: “скорость размножения
бактерий (математически![]() )
)
пропорциональна их числу (n)
в данный момент времени”.
Таким
образом, необходимое дифференциальное
уравнение имеет вид:
 (1)
(1)
где к
— доступный
экспериментальному определения
коэффициент пропорциональности,
зависящий от вида бактерий и параметров
среды их обитания. Дополнительные
данные, необходимые для решения задачи
следуют из начального условия: при t
= 0,
n = n0
, т. е. в начальный момент времени
количество бактерий считается известным
и равным n0
.
Для решения уравнения
(1) произведем разделение переменных и
последующее интегрирование:
 (2)
(2)
Произвольную
постоянную в уравнении (2) удобно
представить в виде lnС .
Из начального условия: C
= n0.
Решая
логарифмическое уравнение (2) с учетом
начального условия, получим искомый
закон изменения числа бактерий со
временем:
![]() .
.
(3)
Произведем
некоторый анализ результата:
1)
Зная коэффициент к и
начальное число бактерий n0 ,
легко определить их число в любой момент
времени t.
2)
Прирост бактериальной массы определяется
через коэффициент к условиями
среды обитания бактерий. Чем больше
значение к,
тем быстрее увеличивается число бактерий

Рис.3
(см.
рис.3). Если существуют факторы,
препятствующие размножению бактерий
(повышенная температура, ионизирующие
излучения и др.), то коэффициент к в
формулах (1) — (3) уменьшается и может
принять отрицательное значение — в этом
случае будет наблюдаться гибель бактерий.
3)
С некоторым риском можно попытаться
придать полученному для бактерий
результату (3) большую общность и
сформулировать утверждение: «любой
биологический вид, находясь в оптимальных
для своего существования условиях,
экспоненциально увеличивает свою
численность со временем». Примеры
справедливости этого утверждения можно
наблюдатьТак, кролики, завезенные в
Австралию, где практически нет хищников,
которые бы ими питались, увеличили свое
число в соответствии с формулой (3) и
стали представлять серьезную опасность
для сельского хозяйства.
2.2. Биомасса
популяции.
Рассмотрим популяцию,
в которой масса особи заметно меняется
в течение жизни, и подсчитаем общую
биомассу популяции.
Пусть ![]() означает
означает
возраст в тех или иных единицах времени,
а N ( ![]() )
)
— число особей популяции, возраст
которых равен ![]() .
.
Пусть, наконец, P ( ![]() )
)
— средняя масса особи возраста ![]() , а М
, а М
( ![]() )
)
— биомасса
всех особей в возрасте от 0 до ![]() .[1]
.[1]
Заметив,
что произведение N( ![]() )
)
P ( ![]() )
)
равно биомассе всех особей возраста ![]() ,
,
рассмотрим разность
M( ![]() +
+
Δ ![]() )
)
– M( ![]() ),
),
где
Δ ![]() >0.
>0.
Очевидно, что эта разность, равная
биомассе всех особей в возрасте
от ![]() до
до ![]() +
+
Δ ![]() ,
,
удовлетворяет неравенствам:
N ( ![]() ) Р (
) Р ( ![]() )Δ
)Δ ![]() ≤
≤
M ( ![]() +
+
Δ ![]() )
)
– M ( ![]() )
)
≤ N( ![]() )P(
)P( ![]() )Δ
)Δ ![]() ,
,
где N ( ![]() ) Р (
) Р ( ![]() )
)
— наименьшее, а — N( ![]() )P(
)P( ![]() )
)
— наибольшее значения функции N
( ![]() )
)
Р ( ![]() ) на
) на
отрезке [ ![]() ,
, ![]() +
+
Δ ![]() ].
].
Учитывая, что Δ ![]() >0,
>0,
из неравенств N ( ![]() ) Р (
) Р ( ![]() )Δ
)Δ ![]() ≤
≤
M ( ![]() +
+
Δ ![]() )
)
– M ( ![]() )
)
≤ N( ![]() )P(
)P( ![]() )Δ
)Δ ![]() ,
,
имеем:
N ( ![]() ) Р (
) Р ( ![]() )
)
≤ ![]() ≤
≤
N( ![]() )P(
)P( ![]() )
)
Из
непрерывности функции N ( ![]() ) Р (
) Р ( ![]() )
)
(ее непрерывность следует из
непрерывности N ( ![]() )
)
и Р ( ![]() )
)
) следует, что
![]() [N (
[N ( ![]() ) Р (
) Р ( ![]() )]
)]
= ![]() [N(
[N( ![]() )P(
)P( ![]() )]
)]
= N
( ![]() )
)
Р ( ![]() )
)
Поэтому
будем иметь:
![]()
![]() = N
= N
( ![]() )
)
Р ( ![]() )
)
или
![]() = N
= N
( ![]() )
)
Р ( ![]() )
)
Следовательно,
биомасса М ( ![]() )
)
является первообразной для N
( ![]() )
)
Р ( ![]() ). Отсюда:
). Отсюда:
M(T)
– M(0) = ![]() N
N
( ![]() )
)
Р ( ![]() )dt
)dt
|
где Т
— максимальный
возраст особи в данной популяции. Так
как М (0),
очевидно, равно нулю, то окончательно
получаем:
М(Т)= ![]() N
N
( ![]() )
)
Р ( ![]() )dt
)dt
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Численность популяции определяется в основном двумя противоположными явлениями — рождаемостью и смертностью. Пусть N — численность популяции. Тогда отношение с/А// означает мгновенную скорость изменения А/, т. е. изменение N в момент времени £, а отношение с1Ы/(М Ы — удельную мгновенную скорость изменения численности популяции. Так, в популяции инфузорий с исходной численностью 100 особей и с численностью 200 особей через час рост популяции равен 100 особей в час, а рост популяции в расчете на одну особь равен 100/100 1 = 1 особь в час.[ …]
Численность популяции — это общее количество особей на данной территории или в данном объеме. Численность популяции зависит от соотношения интенсивности размножения (плодовитости) и смертности. В период размножения происходит рост популяции. Смертность же, наоборот, приводит к сокращению ее численности.[ …]
Под численностью популяции понимают эффективную величину численности, т. е. число размножающихся животных, которое всегда меньше общего числа особей, составляющих популяцию.[ …]
Регуляция численности популяций. Численность любой популяции чрезвычайно динамична и постоянно колеблется вокруг некоторого среднего уровня в соответствии с изменяющимися условиями среды. Выделяют три типа популяционной динамики. Стабильным считается ход численности при изменениях всего в несколько раз, изменчивым — при колебаниях в десятки раз, колебания в сотни и тысячи раз — взрывным. Если вид на время освобождается от действия врагов, возникают взрывы численности (пример, распространение колорадского жука в Ьвропе). У видов, имеющих мало врагов, основную роль в регуляции численности играют внутривидовые связи.[ …]
Мониторинг численности популяций и влияние на них разных факторов. Среди распространенных видов животных наиболее уязвимыми с точки зрения возможности гибели на дороге являются лось, кабан, заяц-беляк, лисица, белка, птицы семейств овсян-никовые и вьюрковые.[ …]
Возрастная структура популяции определяется соотношением особей разного возраста, которое также колеблется. В стабильной популяции рождаемость равна смертности, численность популяции почти не меняется, разновозрастные группы находятся примерно в одинаковом соотношении. В растущих популяциях рождаемость превышает смертность, и численность увеличивается.[ …]
| Регулирование численности популяции (/1) не зависящей от плотно -сти рождаемостью и зависящей от плотности смертностью; (£) зависящей от плотности рождаемостью и не зависящей от плотности смертностью; (В) зависящими от плотности рождаемостью и смертностью. Размер популяции растет, когда рождаемость выше смертности, и падает, когда смертность выше рождаемости. N° — устойчивый равновесный размер популяции, определяемый как интенсивностью не зависящих от плотности процессов, так и интенсивностью и углом наклона графика зависящих от плотности процессов |  |
В природе, однако, рост численности популяций любого вида никогда не бывает бесконечным, поскольку ресурсы, за счет которых существуют виды, на любой территории имеют пределы. Эти пределы называют емкостью среды для конкретных популяций. Например, еловый лес — более емкая среда для белок, нежели смешанный, с березами, поскольку основная пища этих зверьков — семена шишек.[ …]
Периодические колебания численности популяций совершаются обычно в течение одного сезона или нескольких лет. Циклические изменения с подъемом численности в среднем через 4 года зарегистрированы у животных, обитающих в тундре, — леммингов, полярной совы, песца. Сезонные колебания численности характерны и для многих насекомых, мышевидных грызунов, птиц, мелких водных организмов.[ …]
Проблема выживания малых популяций возникла прежде всего как проблема сохранения популяций редких млекопитающих и птиц в зоопарках и питомниках их предварительного размножения для последующего выпуска в природу. Существенным для ее решения и создания моделей выживания малых популяций является понятие эффективной численности (1Че). Она отличается от полной, или действительной, численности популяции (Ы), так как исключает особей, вышедших из репродуктивного возраста й часть молодняка, для которой низка вероятность доживания до этого возраста и оставления потомства. Иными словами, эффективная численность — это численность особей, оставляющих потомство и вносящих свой вклад в генофонд следующего поколения.[ …]
Проблема выживания малых популяций возникла прежде всего как проблема сохранения популяций редких млекопитающих и птиц в зоопарках и питомниках их предварительного размножения для последующего выпуска в природу. Существенным для ее решения и создания моделей выживания малых популяций является понятие эффективной численности (1Че). Она отличается от полной, или действительной, численности популяции (Ы), так как исключает особей, вышедших из репродуктивного возраста й часть молодняка, для которой низка вероятность доживания до этого возраста и оставления потомства. Иными словами, эффективная численность — это численность особей, оставляющих потомство и вносящих свой вклад в генофонд следующего поколения.[ …]
Поддержание определенной численности или равновесное состояние получило название гомеостаза популяций. Рост, снижение или постоянство численности популяций и зависит от соотношения между биотическим потенциалом (прибавлением особей) и сопротивлением среды (гибелью особей), рис. 10.24.[ …]
Важным фактором изменения численности популяций является соотношение полов. Оно редко бывает равным единице, так как в большинстве случаев один из полов преобладает над другим. У позвоночных часто при рождении самцов бывает несколько больше, чем самок. У утиных самцы тоже нередко численно превалируют над самками. Зато в крупных популяциях белок 5сш-ги сагоИпеш1 и 5. т ег доминируют самки.[ …]
Еще одним примером динамики численности популяций являются циклические случаи вспышек массового размножения, вслед за которыми идет резкое сокращение.[ …]
Известно три типа зависимости численности популяции от ее плотности (рис. 7.10). При первом типе (кривая 1) скорость роста популяции уменьшается по мере увеличения плотности. Это широко распространенное явление позволяет понять, почему популяции некоторых животных относительно устойчивы. Во-первых, при увеличении плотности популяции наблюдается снижение рождаемости. Так, в популяции большой синицы при плотности меньше одной пары на 1 га на одно гнездо приходится 14 птенцов; когда же плотность достигает 18 пар на 1 га, выводок составляет менее 8 птенцов. Во-вторых, при увеличении плотности популяции меняется возраст наступления половой зрелости. Например, африканский слон в зависимости от плотности популяции может достигать половой зрелости в возрасте от 12 до 18 лет. Кроме того, этот вид при низкой плотности дает приплод 1 слоненок за 4 года, тогда как при высокой — рождаемость составляет 1 слоненок за 7 лет.[ …]
Эта первая модель, описывающая численность популяции, была предложена Т. Мальтусом еще в конце XVIII в. для описания процесса изменения численности населения. Так как условия неограниченности ресурсов в природе не наблюдаются, или наблюдаются на ограниченных временных интервалах и территориях, модель Мальтуса является приближенной. Т. Мальтус показал ее применимость для анализа динамики популяций колонистов Северной Америки. Закон Мальтуса хорошо описывает динамику искусственных систем, развитие популяций на начальном этапе, пока численность популяций невелика.[ …]
Теперь очевидно, что увеличение численности волков приведет к увеличению численности популяции растений из-за снижения численности оленя. В то же время возрастание биомассы корма оленя вызовет и увеличение численности волков (за счет интенсивного питания и размножения оленей). Однако в таком случае система начинает работать «вразнос» из-за отсутствия механизма отрицательной обратной связи. Фактически же увеличение биомассы растений не может быть беспредельным: оно всецело зависит от абиотической компоненты среды — почвенных условий, прихода солнечной энергии и ассимиляции питательных веществ продуцентами, каковыми и являются растения.[ …]
Количественным показателем оценки популяции является плотность — численность популяции, отнесенная к единице занимаемого пространства. Для характеристики пространственного распределения особей применяют методы математической статистики, которые позволяют оценить дисперсию наблюдаемого распределения плотности и сопоставить ее со средним значением плотности. При случайном распределении дисперсия равна среднему значению: а2 = т, при регулярном — дисперсия меньше среднего: ст2 < т и при пятнистом — дисперсия больше сред-■ него: ст2 > от. В соответствии с этим отношение дисперсии к среднему значению есть показатель степени пространственной агрегированности. Если величина этого показателя примерно равна единице — распределение случайное; меньше единицы — регулярное; больше единицы — пятнистое.[ …]
Рождаемость и смертность регулируют численность популяции и ее возрастной состав.[ …]
Поскольку длительность существования популяции значительно превышает продолжительность жизни отдельных особей, в ней всегда присутствует смена поколений. И даже если численность популяции постоянна, то постоянство является результатом динамического равновесия прибыли и убыли особей.[ …]
Самый бесхитростный способ определения численности популяции— это простой подсчет всех имеющихся особей. Так можно поступить со многими растениями; можно пересчитать и животных— если это животные сидячие или крайне малоподвижные или если они достаточно крупные, хорошо заметные и легко различимые для того, чтобы можно было «просмотреть» всю популяцию, никого не упустив и никого не сосчитав дважды. Но и тогда, когда такие «поголовные переписи» осуществимы, они зачастую требуют больших затрат времени и квалифицированного труда.[ …]
В этом и состоит принцип минимального размера популяций. Минимальная численность популяций, обеспечивающая существование вида, является специфической для разных видов. Выход за пределы минимума грозит для популяции гибелью. Дальнейшее сокращение, например, тигра на Дальнем Востоке, неизбежно приведет к их автоматическому вымиранию из-за того, что оставшиеся единицы, не находя с достаточной частотой партнеров для размножения, вымрут на протяжении немногих поколений. В таком же положении могут оказаться и редкие растения, такие, как орхидея «венерин башмачок» и другие.[ …]
Это уравнение представляет собой модель роста популяции, ограниченного внутривидовой конкуренцией. Суть этой модели в том, что нереалистичная константа Я в уравнении (6.1) заменена на фактическую чистую скорость воспроизводства, /?/(1 — -a-Nt), которая уменьшается по мере роста численности популяции (А/’ ).[ …]
Точка В, напротив, отражает ситуацию, в которой численность популяции (А ) гораздо выше и где в значительной степени существует внутривидовая конкуренция. В самом деле, в точке В фактическая чистая скорость воспроизводства в результате конкуренции настолько снижена, что популяция в целом может не более, чем восстанавливать в каждом поколении свою численность, потому что количество родившихся особей уравновешивается количеством погибших. Другими словами, ЛГ(+1 в точности соответствует и А /Л/ж равно 1, а численность популяции, при которой имеет место это соотношение, называется предельной плотностью насыщения К (рис. 6.5).[ …]
На графиках А и Б показаны изменения во времени численности популяций в неустойчивой (А) и устойчивой (£) ситуациях, / — хозяин; // — паразит. Графики В и Г представляют собой «фазовые диаграммы», показывающие изменения во времени размеров популяций относительно друг друга. Стрелками указано направление изменений во времени.[ …]
Совокупность всех факторов, способствующих увеличению численности популяции, называется биотическим потенциалом. Несмотря на то, что у разных видов составляющие биотического потенциала неодинаковы, имеется общее свойство. У всех видов он достаточно высок для стремительного увеличения численности при благоприятных условиях среды. Рост популяции может быть столь быстрым, что может привести к популяционному взрыву. Однако следует отметить, что повышение плотности популяций сверх оптимальной оказывает на них неблагоприятное воздействие, так как при этом иссякает кормовая база, сокращается жизненное пространство, появляются эпизоотии и т. д. (рис. 10.22).[ …]
ДЕПРЕССИЯ — 1. Состояние угнетения, в котором находится популяция в неблагоприятный период жизни. Сопровождается сокращением численности популяции (депопуляцией). 2. Д. подземных вод — снижение свободной или напорной поверхности подземных вод, часто приводит к их истощению и загрязнению.[ …]
Нашим следующим шагом будет вывод общего соотношения, связывающего численность популяции со скоростью ее увеличения и временем; соотношение это будет таким, что единицей измерения времени вовсе не обязательно должно будет служить поколение. Вообразим популяцию с начальной численностью в 10 особей; пусть, далее, по прошествии последовательных промежутков времени численность ее возрастает сначала до 20, затем до 40, 80, 160 особей, и т. д. Начальную численность популяции мы обозначим через Ы0 (имея в виду число особей в тот момент, когда начинается отсчет времени); численность той же популяции по истечении одного промежутка времени — через Л ь по истечении двух промежутков — через Ы2, а по истечении произвольного числа промежутков (¿)—через Nt. В нашем примере Л 0=10, а Л 1 = 20. Можно еще написать, что Л = М0/?, где (в нашем примере равное 2) —показатель, именуемый «основной чистой скоростью воспроизводства» либо «основной чистой удельной скоростью увеличения популяции». Когда /?>1, численность популяции будет, очевидно, нарастать, а когда Я<С1— снижаться.[ …]
| Зависимые от плотности рождаемость и смертность обусловливают регуляцию численности популяции. Если оба (Л) или один из параметров (Б и В) зависят от плотности, то кривые пересекаются. Плотность, при которой происходит пересечение, называется предельной плотностью насыщения (/(). При плотности ниже К популяция растет, при плотности выше К — уменьшается; при плотности, равной К, наблюдается устойчивое равновесие. На рисунке дана лишь самая общая схема | 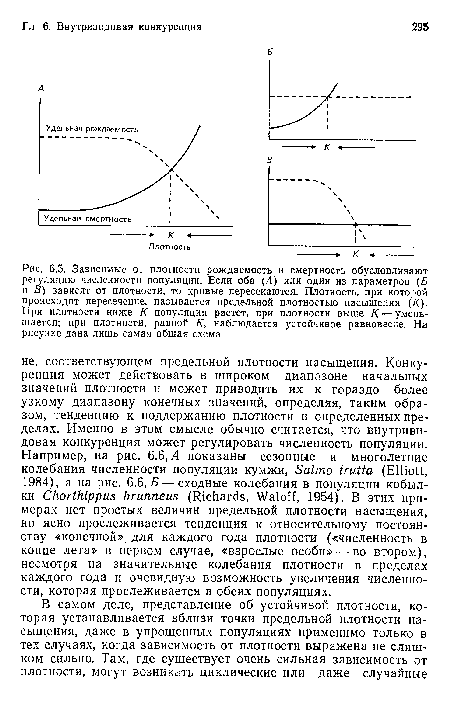 |
Второе обобщение дополняет первое и носит название теории биоценотической регуляции численности популяции К. Фри-дерикса (1927): регуляция численности популяции есть результат комплекса воздействий абиотической и биотической среды в местообитании вида.[ …]
Джерасси и сотр. [460] считают, что экономические и научные проблемы, связанные с необходимостью регулировать численность популяций насекомых, во многих отношениях напоминают проблему регуляции народонаселения.[ …]
Другим важным аспектом в методическом отношении может иметь значение так называемого «эффекта нагрузки» и необходимая исходная численность популяции в опыте, не оказывающая влияния на дальнейшее развитие всей культуры. В литературе вообще известно много фактов, подтверждающих непосредственное влияние количества исходных особей или посевного материала (как на уменьшение, так и на увеличение их) на скорость и характер их дальнейшего развития (Мейнелл, Мейнелл, 1967; Ивлева, 1969).[ …]
В этих формулах В и — коэффициенты миграции особей в пределах ареала распространения. Если ареал ограничен, качественная картина изменения численности популяций сохраняется. Однако если ареал не ограничен, то возможно возникновение решений типа волн. Важным свойством пространственного поведения системы хищник-жертва является возникновение пространственных неоднородностей плотности распределения особей — так называемых диссипативных структур. Таким образом, если в точечных моделях наблюдаются автоколебательные режимы, то в пространственно распределенных системах имеют место диссипативные структуры.[ …]
Когда началось вторжение европейцев в Северную Америку, в регионе, который впоследствии станет Восточной Канадой и северо-восточными Соединенными Штатами, постоянно обреталась популяция моржей, насчитывавшая не менее чем три четверти миллиона особей. По крайней мере еще четверть миллиона обитала в прилежащих морях к северу. К 1972 году общая численность популяции моржей восточной части Северной Америки составляла от 5 до 10 тысяч особей, причем их пребывание было ограничено арктическими и субарктическими водами. Несмотря на официальную защиту, их численность все еще сокращается, прежде всего из-за их клыков, которые теперь снова вошли в моду как в виде дорогих сувениров, так и в качестве сырья для резьбы по кости. В 1981 году много тонн нелегально вывезенной североамериканской моржовой кости поступило на международные рынки, где было продано по цене до 150 долларов за фунт. Агенты Федерального управления по защите животных США на Аляске конфисковали 10 тысяч фунтов клыков за один день. Такое большое количество кости потребовало уничтожения как минимум 750 взрослых моржей. Именно такое число обезглавленных моржей недавно было выброшено океаном на побережье Сибири напротив Аляски, и Советский Союз подал официальный протест государственному департаменту США с осуждением имевшей место резни.[ …]
С экологической точки зрения загрязнение поступает в экосистему. Поступление загрязнений изменяет функционирование экосистемы. Изменяются потоки энергии и вещества, продуктивность, численность популяций и т. п. Загрязнение природной среды может происходить от естественных источников (наводнение, извержение вулкана, падение метеорита и т.д.) и в рёзультате человеческой деятельности. Таким образом, следует различать природные и антропогенные загрязнения. Загрязнение может иметь физическую, химическую, биологическую природу.[ …]
Таким образом, в биоценозе между организмами наблюдаются самые разнообразные формы взаимоотношений, построенные на пищевых, пространственных и других типах взаимодействия, благодаря которым регулируется численность популяций и повышается устойчивость сообщества.[ …]
Борьба с мухой каллитрогой, опасным вредителем домашнего скота, с помощью выращивания в искусственных условиях насекомых, у которых облучением вызывается стерильность, открывает новые возможности биологической борьбы. Обычно самки моногамны и спариваются только один раз. После спаривания с облученной мужской особью они откладывают только стерильные яйца. Основой борьбы является относительно низкая численность популяции в зимний период. Если число выпускаемых облученных самцов будет постоянно превышать численность нормальных,то число нормальных яиц будет непрерывно снижаться (независимо от того, что инстинкт спаривания у самок будет сохраняться). Чем больше будет доля облученных самцов по сравнению с нормальными, тем успешнее будет борьба. Полученные результаты позволяют предполагать, что этот способ перспективен для борьбы с вредителями растений1.[ …]
Был предпринят целый ряд попыток проанализировать математически зависимость между сложностью сообщества и его устойчивостью, в большинстве из которых авторы пришли к примерно одинаковым выводам. Обзор таких публикаций дал Мей (May, 1981). В качестве примера рассмотрим его работу (May, 1972), демонстрирующую как сам метод, так и его недостатки. На каждый вид влияли его взаимодействия со всеми остальными видами; количественно влияние плотности вида / на рост численности i оценивалось показателем р . При полном отсутствии влияния равен нулю, у двух конкурирующих видов Рц и Pji отрицательны, в случае хищника (¿) и жертвы (/) Ру положителен, a jjji — отрицателен.[ …]
Модель есть абстрактное описание того или иного явления реального мира, позволяющее делать предсказания относительно этого явления. В простейшем случае модель может быть словесной или графической (т. е. неформализованной). Однако если мы хотим получить удовлетворительные количественные прогнозы, то модель должна быть статистической и строго математической (т. е. формализованной). Например, математическое выражение, описывающее изменения численности популяции насекомых и позволяющее предсказывать эту численность в определенные моменты времени, следует считать моделью, полезной с биологической точки зрения. А если рассматриваемую популяцию составляет вид-вредитель, то эта модель приобретает еще и экономическое значение.[ …]
Для управления экосистемами не требуется регуляция извне — это саморегулирующаяся система. Саморегулирующий гомеостаз на экосистемном уровне обеспечен множеством управляющих механизмов. Один из них — субсистема «хищник—жертва» (рис. 5.3). Между условно выделенными кибернетическими блоками управление осуществляется посредством положительных и отрицательных связей. Положительная обратная связь «усиливает отклонение», например увеличивает чрезмерно популяцию жертвы. Отрицательная обратная связь «уменьшает отклонение», например, ограничивает рост популяции жертвы за счет увеличения численности популяции Хищников. Эта кибернетическая схема (рис. 5.3а) отлично иллюстрирует процесс коэволюции в системе «хищник—жертва», так как в этой «связке» развиваются и взаимные адаптационные процессы (см. рис. 3.5). Если в эту систему яе вмешиваются другие факторы (например, человек.уничтожил хищника), то результат саморегуляции будет описываться гомеостатическим плато (рис. 5.3 б) — областью отрицательных связей, а при нарушении системы начинают преобладать обратные положительные связи, что может привести к гибели системы.[ …]
Вместе с тем нельзя ожидать, что рождаемость, скорость роста и выживаемость консументов по мере увеличения доступности пищи будут расти до бесконечности. Консументы достигают состояния насыщения, и скорость потребления пищи постепенно выходит на постоянный уровень, при котором она не зависит от количества доступного ¡корма (рис. 8.7); следовательно, выигрыш, получаемый консументом, также достигает постоянного уровня. Таким образом, существует предельное количество пищи, ¡которое данная популяция консумента в состоянии съесть, предел вредного воздействия на популяцию своей жертвы и предел, до которого может увеличиваться численность популяции консумента.[ …]
Подобно всем другим областям знания, экология развивалась непрерывно, но неравномерно. Труды Гиппократа, Аристотеля и иных древнегреческих философов содержат сведения явно экологического характера. Вместе с тем греки не знали слова «экология». Многие великие деятели «биологического возрождения» (XVIII— XIX вв.) внесли свой вклад в эту область. Например, Антон ван Левенгук, более известный как один из первых микроскопистов начала XVIII в., был также пионером в изучении «пищевых цепей» и динамики численности популяций — двух важных разделов современной экологии.[ …]
Подобно всем другим областям знания экология развивалась непрерывно, но неравномерно. Труды Гиппократа, Аристотеля и других древнегреческих философов содержат сведения явно экологического характера. Однако греки не знали слова «экология». Термин этот недавнего происхождения. Он был предложен немецким биологом Эрнстом Геккелем в 1869 г. Многие великие деятели «биологического Возрождения» (XVIII—XIX вв.) внесли свой вклад в эту область, хотя название «экология» долгое время не употреблялось. Например, Антон ван Левенгук, более известный как один из первых микроскрпистов начала XVIII в., был также пионером в изучении «пищевых цепей» и регулирования численности популяций (Эгертон, 1968) — двух важных разделов современной экологии. Как самостоятельная наука экология сформировалась приблизительно к 1900 г., но лишь в последнее десятилетие это слово приобрело особую популярность. В наши дни каждый остро осознает важность наук о среде для поддержания и повышения уровня современной цивилизации. Экология быстро становится отраслью науки, теснейшим образом связанной с повседневной жизнью каждого человека, будь то мужчина, женщина или ребенок.[ …]
Численность — это общее количество особей одного вида на участке ареала с однородными экологическими условиями. Например, дальневосточная популяция тигра насчитывает около 600 особей, ладожской нерпы — около 10 тыс. особей, азиатского льва — более 600 особей, популяция зубра на территории Национального парка Беловежская пуща — более 500 особей. Определить общую численность популяции путем прямого подсчета можно только для крупных и хорошо заметных особей. Численность популяции постоянно изменяется. Но рост численности популяций любого вида в природе никогда не бывает бесконечным, потому что ресурсы, за счет которых существуют виды (пища, убежища, подходящие места для размножения), на любой территории имеют пределы. В то же время численность популяций не может снижаться бесконечно, так как для обеспечения воспроизводства потомства требуется наличие определенного числа особей. Следовательно, колебания численности ограничиваются верхним и нижним пределами. Выход за эти пределы может привести к гибели популяции.
Верхний предел численности — максимальное количество особей популяции, потребности которых могут быть удовлетворены ресурсами данного местообитания без заметного его нарушения. Он зависит от количества корма, площади занимаемой территории, воздействия экологических факторов и в экологии называется емкостью среды. Однако популяция при быстром росте легко может превысить емкость среды, и тогда начинается гибель особей из-за нехватки корма. Из-за повышенной контактности возникают эпидемии, что в конечном итоге может привести к гибели всей популяции.
Нижний предел численности — минимальное количество особей, способных обеспечить воспроизводство и длительное существование популяции. Он зависит от биологических свойств организмов и является величиной постоянной для всех популяций в пределах вида. Снижение численности ниже нижнего предела является причиной снижения возможности встречи полов для размножения особей. Это неизбежно приводит к вымиранию популяции. Вот почему популяции с очень малой численностью особей длительно существовать не могут. Известны случаи вымирания популяций даже в заповедниках со строгим режимом охраны, когда их численность снижалась ниже нижнего предела.
Плотность — количество особей популяции в расчете на единицу площади (м2, га, км2) или объема (л, м3, км3). Например, 150 сосен на 1 га соснового бора, 800 клеток хлореллы в 1 л воды характеризуют плотность популяций этих видов. Плотность прямо пропорциональна численности, поэтому показатель численности чаще всего рассчитывают путем умножения плотности на площадь ареала популяции. При увеличении численности плотность не повышается или даже снижается лишь в том случае, если возможно расселение особей вследствие расширения ареала. При отсутствии расселения показатель плотности популяции позволяет количественно сравнивать отдельные популяции независимо от общего размера занимаемой ими территории или акватории.
Различают два вида плотности. Средняя плотность — количество особей в расчете на единицу площади всей занимаемой территории. Экологическая плотность — количество особей в расчете на единицу площади территории, пригодной для обитания. При неравномерном заселении особями территории эти показатели могут существенно различаться.
Плотность, как и численность, имеет верхний и нижний пределы, выход за которые может приводить к гибели популяции. Поэтому прогноз для будущего существования популяции можно давать и на основании сравнения плотности с ее пределами.
Следовательно, если численность или плотность близки к верхнему пределу, то часть особей следует изъять из популяции. Это может быть переселение на свободную территорию, использование в хозяйственных целях, санитарный отстрел животных или вырубка деревьев. Если же численность или плотность приближаются к нижнему пределу, то следует осуществлять охрану популяции или вселять новые особи, чтобы не допустить снижения численности ниже нижнего предела.
Повторим главное. Популяция — способная к саморегуляции группа особей одного вида, длительно обитающих на общей территории, свободно скрещивающихся между собой и дающих плодовитое потомство. Любая популяция имеет биологические и групповые признаки (свойства и структуру). Свойства популяции разделяют на две группы: статические и динамические. К статическим свойствам относятся численность и плотность. Диапазон изменения численности и плотности ограничивают верхний и нижний пределы, выход за которые приводит к гибели популяции. Для популяции можно рассчитать среднюю и экологическую плотность.