Основными
параметрами популяции, прежде всего,
являются ее численность
и плотность.
Численность
популяции – это
общее количество особей на данной
территории или в данном объеме.
Численность
никогда не бывает постоянной и зависит
от соотношения интенсивности размножения
(плодовитости) и смертности. В процессе
размножения происходит рост популяции,
смертность же приводит к сокращению ее
численности.
Плотность
популяции определяется количеством
особей либо биомассой на единицу площади
или объема.
Например,
150 растений сосны на 1 га или 0,5 г циклопов
в 1 м3
воды характеризуют плотность популяции
этих видов. Плотность популяции также
изменчива
и зависит от численности. При
возрастании численности плотность не
увеличивается лишь в том случае, если
возможно расселение популяции,
расширение ее ареала. Любой популяции
свойственна тенденция к расселению.
Расселение продолжается до тех пор,
пока популяция не встретится с какой-либо
преградой.
В
связи с размерами особей того или иного
вида и, следовательно, ареалов его
популяций варьируется и численность
особей в каждой из них.
У насекомых, мелких травянистых растений
она
может достигать сотни тысяч и более.
Популяции же крупных животных и больших
древесных
растений чаще всего
бывают
небольшими по численности.
Но так как размеры ареала популяции
непостоянны, то и численность особей,
ее слагающих, претерпевает различные
изменения. Нередко эти изменения связаны
с деятельностью человека. Но основными
причинами их являются изменения условий
существования. Как известно, динамика
популяций белок, зайцев, многих птиц в
большинстве случаев зависит от урожая
кормов. Иногда причины динамики популяций
не ясны. Но в природе не отмечено ни
одной популяции, которая бы оставалась
стабильной даже на протяжении короткого
отрезка времени. Это связано, прежде
всего, с воздействием абиотических
факторов на интенсивность роста и
размножения, на смертность, скорость
развития. Немалую роль в данном
процессе играют уничтожение хищниками
жертв, конкурентные взаимоотношения,
влияние паразитов на жизнедеятельность
хозяев и т. д.
Однако,
несмотря на изменчивость, численность
популяции колеблется не беспредельно.
Популяция, как и любая биологическая
система, обладает способностью к
саморегуляции. Для популяции каждого
вида всегда имеются верхние и нижние
пределы плотности, за границы которых
она выходить не может. При благоприятном
сочетании всех условий плотность
популяции удерживается на каком-то
оптимальном уровне, незначительно
отклоняясь от него в ту или другую
сторону. Такие колебания численности
обычно носят правильный, регулярный
характер и четко отражают реакцию
популяции на конкретные условия среды.
Тем не менее, сезонные колебания
численности, особенно у мелких животных
(мышевидные грызуны, насекомые, некоторые
птицы), могут быть весьма значительны.
Так, численность мышевидных грызунов
в течение одного сезона иногда
увеличивается в 300 — 500 раз, а некоторых
насекомых в 1300 — 1500 раз. Причины вспышек
массового размножения бывают самыми
различными. Они, как правило, сменяются
резким сокращением численности.
Падение
плотности ниже оптимальной приводит
к ослаблению защитных реакций популяции,
снижает ее плодовитость и вызывает ряд
других отрицательных явлений. В популяциях
с минимальной численностью нередко
складываются весьма сложные ситуации.
Изменяющиеся совокупности ее легко
могут быть сведены к нулю. Это неизбежно
приводит к вымиранию популяции. У
двуполых видов, например, достаточно,
чтобы до нуля сварьировал один из полов.
Вот почему популяции с очень малой
численностью особей длительно
существовать не могут. Такие случаи
известны даже в заповедниках со
строгим режимом.
Повышение
плотности сверх оптимальной также
неблагоприятно сказывается на популяции,
поскольку при этом иссякает кормовая
база, сокращается жизненное пространство,
появляются эпизоотии и т. д.
Соседние файлы в папке Биосфера_Экосистемы
- #
- #
- #
- #
- #
- #
- #
А.С. Степановских
Экология. Учебник для вузов
М.: ЮНИТИ-ДАНА, 2001. — 703 с.
Содержание статьи:
- 1 9. Структура и динамика популяций
- 1.1 9.3. Численность и плотность популяций
9. Структура и динамика популяций
9.3. Численность и плотность популяций
Основными показателями структуры популяций является численность и распределение организмов в пространстве и соотношение разнокачественных особей. В связи с размерами ареала популяций может значительно изменяться и численность особей в популяциях.
Численность популяции — это общее количество особей на данной территории или в данном объеме. Зависит от соотношения интенсивности размножения (плодовитости) и смертности. В период размножения происходит рост популяции. Смертность же, наоборот, приводит к сокращению ее численности.
Плотность популяции определяется количеством особей или биомассой на единицу площади либо объема, например: 400 деревьев на 1 га, 0,5 г циклопов в 1 м3 воды. Нередко важно различать среднюю плотность, т. е. численность или биомассу на единицу всего пространства, кудельную или экологическую плотность — численность или биомассу на единицу обитаемого пространства, доступной площади объема, которые фактически могут быть заняты популяцией.
Плотность популяции отличается изменчивостью и зависит от ее численности. При возрастании численности не наблюдается увеличение плотности лишь в том случае, когда возможно распределение популяции, расширение ее ареала. Особи, составляющие популяции, имеют различные типы пространственного распределения, выражающие их реакции на различные влияния, например, добычу и благоприятные физические условия или конкурентные реакции. Различают три типа распределения или расселения особей внутри популяции: равномерное, случайное и групповое (рис. 9.2).
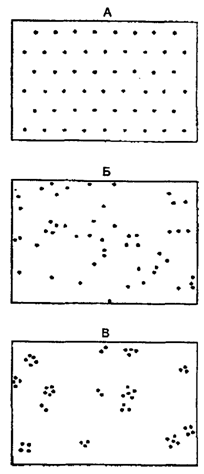
Рис. 9.2. Основные типы распределения особей в популяции:
А — равномерное распределение; Б – случайное распределение; В — групповое распределение (по Одуму, 1986)
Равномерное распределение в природе чаще связано с острой конкуренцией между разными особями. Такой тип распределения отмечают у хищных рыб и у колюшек с их территориальным инстинктом и сугубо индивидуальным характером.
Случайное распределение имеет место только в однородной среде. Так на первых порах распределяется тля на поле. По мере ее размножения распределение приобретает групповой или пятнистый (конгрегационный) характер.
Групповое распределение встречается наиболее часто. Так, в сосновом лесу деревья вначале расселяются группами, а в дальнейшем их размещение становится равномерным. Популяции групповое распределение обеспечивает (по Ю. Одуму, 1986) более высокую устойчивость по отношению к неблагоприятным условиям по сравнению с отдельной особью. Животные, ведущие подвижный образ жизни, как правило, распределяются активно, что приводит к интенсивному перемешиванию популяций и стиранию границ между ними. Например, очень подвижные и активно перемещающиеся песцы, другие животные, птицы имеют огромные ареалы без резких границ между популяциями.
У пассивно передвигающихся и малоподвижных организмов, наоборот, популяции четко разграничены даже на относительно небольшой территории. Таковы популяции наземных моллюсков, многих земноводных. Размеры ареала популяции зависят от величины особей, составляющей ее. Мелкие особи занимают сравнительно небольшие ареалы, тогда как у видов с крупными особями они обширны. Вместе с тем это правило имеет много исключений. Так, территория, занимаемая популяцией прыткой ящерицы Lacerta agilis, может колебаться от 0,1 до нескольких гектаров.
Знание типа распределения организмов имеет большое значение при оценке плотности популяции методом выборки (в случае группового размещения площадь выборки должна быть большая). Возьмем n выборок. Среднее число особей в каждой выборке обозначим через m и получим рассеяние или дисперсию S2 по формуле:
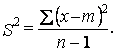 (9.1)
(9.1)
При равномерном распределении дисперсия S2 равна нулю, так как число особей в каждой выборке постоянно и равно среднему. В случае случайного распределения среднее m и дисперсия S2 равны. При групповом распределении рассеяние S2 выше среднего и разница между ними тем больше, чем сильнее тенденция животных к образованию скоплений.
Плотность популяций
2014-06-01
С численностью популяции неразрывно связана ее плотность. Под плотностью (плотностью — у растений) понимают среднее количество особей (или биомассу), приходящаяся на единицу площади или объема (на 1 м2, на 1 га, в 1 мм2 и т.д.). Не надо путать понятия численность и плотность. Для примера: численность популяции бука на всей ее территории составляет 1,5 тыс. деревьев при плотности 150 деревьев на 1 га; плотность рыбы — 200 кг на 1 га поверхности пруда; 5 млн. особей хлореллы в 1м3 воды.
Отдельно следует выделить понятие минимальная, максимальная, средняя и экологическая плотность популяции.
Минимальная плотность особей на данной территории не может обеспечить дальнейший рост популяции, поскольку не создает необходимых условий для их размножения и возможностей для существования популяции в пределах определенной экосистемы.
Максимальная плотность — такое количество особей, которую экосистема не может поддерживать из-за недостатка территории и кормовых ресурсов.
Средняя плотность — среднее число особей (биомассы), приходящейся на единицу (объем) всей территории популяции.
Плотность определяет насыщенность территории представителями данного вида. Особенно важно знать, увеличивается или уменьшается плотность, определяющая направление развития популяции, от чего зависит ее существование.
Итак, понятие численность и плотность популяции четко разграничены: 100 деревьев могут расти на площади 1 га — это невысокая плотность; 100 деревьев на 0,5 га — это вдвое больше плотность.
С увеличением численности плотность популяции возрастает. В случае, когда есть свободная территория для расселения, плотность может даже снижаться. Такое явление в природе наблюдается часто, каждое живое существо пытается выжить и улучшить условия своего существования, а каждый вид — расширить свой ареал.
Поскольку состав популяции определяется не случайной комбинацией особей, а определенными взаимосвязями вида с окружающей средой, определяют также экологическую плотность. Она выражается числом особей (или биомассой) на единицу доступной для заселения части территории (или объема).
Средняя и экологическая плотность тождественны и характеризуют различные состояния популяции. С изменением размеров территории популяции при одной и той же численности особей, плотность его изменяется. На основе сравнения средней и экологической плотности рыб в связи с изменением условий среды и циклом размножения аиста в национальном парке Эверглейдс (Флорида) Коль (1964) установил, что средняя плотность мелких рыб со снижением уровня воды уменьшается. Экологическая плотность их возрастает, поскольку сократилась площадь водного зеркала,
и число рыб на единицу водной поверхности водоема увеличивается. Аист выступает как хищник, питается рыбой. Поэтому он приурочивает откладывания яиц до времени высокой экологической плотности рыб, чтобы облегчить кормление птенцов.
При изучении популяционной плотности необходимо учитывать экологическое состояние ее среды на момент исследования.
От плотности популяции зависит влияние ее на экосистему, независимо от того, из каких представителей состоит популяция. Если один дикий кабан, зубр, лось приходится на несколько гектаров леса, это не нанесет ему существенный вред. Несколько особей этих видов на 1 га являются нежелательными.
Плотность популяции очень изменчива, но ее рост ограничен. Существуют верхние и нижние границы числа особей, обусловленные размерами живых существ, их образом жизни и резервом кормов. Например, в лесу может жить в среднем 100 птиц на 1 га, 20 тыс. грунтовых членистоногих на 1 м2 и только 10 дождевых червей на 1 га. Верхний предел плотности определяется трофическими связями и потоком энергии в экосистеме. Нижняя граница плотности составляет такое минимальное количество особей определенного вида, которая обеспечивает гомеостаз в стабильных экосистемах и возможность существования в ней данной популяции.
Следовательно, важно при изучении популяции в определенных экосистемах (биогеоценозах) учитывать их максимальную и минимальную плотности.
Точно определить плотность популяции трудно в связи с тем, что особи на ее территории часто распределены отдельными группами, неравномерно.
Это одинаково касается растений и животных. Что касается последних, то значительную трудность представляет и тот факт, что животным присущ подвижный образ жизни и миграция из одной популяции в другую.
В таких случаях определяют относительную плотность. Показатели относительной плотности широко используют для популяций крупных животных и наземных растений. Такие данные необходимы для ежегодного регулирования охоты на водоплавающих птиц, промысловых зверей, возможностей и норм лесозаготовки и т.д..
ПЕДАГОГИЧЕСКИЙ УНИВЕРСИТЕТ
«ПЕРВОГО СЕНТЯБРЯ»
ЧЕРНОВА Н.М.
Курс «Учителю биологии об основах
экологии»
Учебный план курса
|
№ газеты |
Учебный материал |
|
17 |
Лекция 1. Почему надо изучать |
|
18 |
Лекция 2. Организм и среда |
|
19 |
Лекция 3. Экологические адаптации |
|
20 |
Лекция 4. Средообразующая роль |
|
21 |
Лекция 5. Биоценозы |
|
22 |
Лекция 6. Популяции |
|
23 |
Лекция 7. Экосистемы |
|
24 |
Лекция 8. Биосфера |
|
Итоговая работа. |
Лекция 6. Популяции
Популяция – это население вида на
определенной территории. Всё географическое
пространство, в пределах которого встречается
вид, носит название ареал. Любой ареал
неоднороден, особенно если он велик и обширен.
Поэтому вид в общих границах своего
распространения распределен неравномерно, и в
его пространственной структуре выделяются
отдельные популяции, населяющие более или менее
обособленные территории. И степень обособления
отдельных популяций, и занимаемые ими территории
или акватории чрезвычайно различны в связи с
разнообразием и особенностями видов. У крупных
подвижных животных, например волков или лосей, мы
выделяем популяции на больших территориях,
обособляемых какими-либо географическими
границами (например, большими реками, хребтами и
т.п.), тогда как для мелких мышевидных грызунов
особенности населения могут различаться в
пределах соседних биотопов (ельник, березняк,
разнотравный луг и др.). Некоторые виды
распространены резко мозаично (например, окуни в
разных озерах), у других четкую границу между
популяциями провести трудно, вид распространен
почти повсеместно, но на разных территориях его
численность различна.
Численность – основная
характеристика популяций. Данные о численности в
разных местах расширяют и уточняют наши
представления об адаптированности вида к тем или
иным условиям среды.
Любая популяция – это не просто сумма
особей одного вида. Это сложная надорганизменная
система с внутренними связями и свойствами,
которые не присущи отдельным составляющим ее
организмам.
Что связывает друг с другом
представителей одного вида, населяющих общую
территорию? Прежде всего, это отношения,
связанные с воспроизводством: между
представителями разного пола, между
родительским и дочерними поколениями. Не менее
важна вторая категория отношений –
экологические. Они обеспечивают распределение
ресурсов. В популяциях разных видов встречаются
все типы биотических связей, которые существуют
и между представителями разных видов, но
наиболее развиты два прямо противоположных
варианта: конкуренция и мутуализм. Переплетаясь
друг с другом, эти отношения обеспечивают
устойчивое существование и воспроизведение вида
в конкретных местообитаниях.
Любая популяция очень динамична, в ней
постоянно происходят изменения: гибнут от
врагов, старости или непогоды отдельные особи,
рождаются новые, меняется возрастной состав,
происходит расселение или вселение
представителей вида с других территорий. Все эти
процессы отражаются в динамических
характеристиках популяций: темп роста или
убыли численности, рождаемость, смертность,
темпы выселения или вселения (эмиграции и
иммиграции). В динамических показателях всегда
учитывается время, в течение которого происходят
те или иные изменения. Если же мы хотим
охарактеризовать состояние популяции в
какой-либо определенный момент времени, то
прибегаем к статическим характеристикам:
численность, плотность, возрастная и половая
структура, распределение в пространстве и т.п. На
динамику популяций большое влияние оказывает
окружающая среда, т.к. виды существуют в
постоянно изменяющихся условиях.
Численность популяции, т.е.
суммарное количество особей, определить чаще
всего бывает нелегко. Можно пересчитать,
например, всех амурских тигров, но сложнее – всех
окуней в большом озере, всех мух в окрестностях
скотного двора или всех дождевых червей в почве
луга. Поэтому в количественной оценке популяций
чаще используют такой показатель, как плотность
– число особей, приходящихся на определенную,
условно выбранную, единицу пространства, где их
можно с достаточной точностью подсчитать.
Плостность популяции дождевых червей, таким
образом, – это их число под 1 м2 поверхности почвы;
плотность популяции зябликов в лесу – число
замеченных птиц на 1 км учетного маршрута; рачков
дафний в пруду – их обилие в 1 дм3 воды на
определенной глубине и т.п. От плотности
популяций существенно зависит и интенсивность
внутрипопуляционных отношений.
Описание возрастной и половой
структуры популяции носит название демография (от
греч. слов «демос» – народ и «графос» – пишу,
описываю). Необходимость и значение
демографических исследований были поняты
сначала применительно к человеку. Еще в XVI в. было
подробно учтено соотношение жителей Лондона по
полу и возрасту, на основании чего оценена
скорость роста населения. Это было очень важно
для планирования действий по обеспечению
жителей продовольствием и водой и созданию
соответствующих условий жизни. В современной
службе городов строгий учет демографических
характеристик населения – необходимое условие
для решения основных социальных вопросов:
строительства жилья и детских учреждений,
пенсионного обеспечения, планирование населения
и т.п.
В природных условиях демографические
особенности любой популяции во многом
определяют ее ближайшее будущее: возможность
размножения и способность сопротивления
действию неблагоприятных факторов. На разных
стадиях развития организмов требования к среде
изменяются и иногда очень значительно. Может
происходить смена сред обитания, изменение типа
питания, характера передвижения, активности,
выносливости к различным погодным условиям и т.п.
Особенно резко это выражено у животных с
метаморфозом. Достаточно вспомнить гусениц и
бабочек чешуекрылых, личинок и имаго майских
жуков, головастиков и взрослых лягушек и т.п. Но и
у организмов с прямым развитием характер
отношений со средой очень сильно зависит от
возраста. Как правило, молодые и стареющие особи
гораздо чувствительнее к переменам условий.
Популяции, в которых преобладают эти возрастные
группы, могут резко снижать численность в
неблагоприятные периоды.
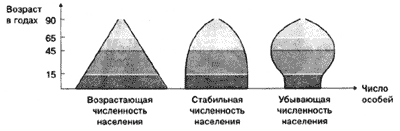
Примеры возрастной структуры
популяции человека
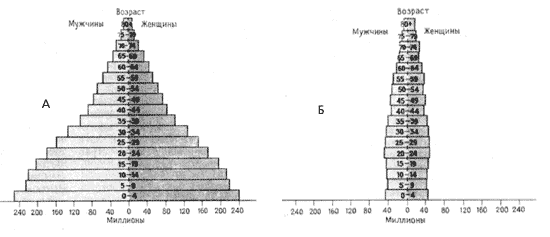
Половозрастные пирамиды: А – для
развивающихся стран; Б – для развитых стран
Половая и возрастная структура
популяций зависит от двух главных причин:
биологических особенностей вида и конкретных
экологических условий его существования. Для
многих видов характерна раздельнополость, и
популяции состоят соответственно из самцов и
самок. Однако есть множество видов, у которых
каждая особь гермафродитна, т.е. имеет и женскую,
и мужскую половые системы (например, дождевые
черви, брюхоногие моллюски, некоторые рыбы, ряд
ракообразных). У таких видов экологические
условия часто определяют, какая из половых
систем будет функционировать нормально, и какое
реальное соотношение самцов и самок окажется в
популяции. Но и при генетическом разделении
полов соотношение 1:1 редко выдерживается.
Экологические различия самцов и самок часто
приводят к разной их смертности в течение жизни.
Например, у антарктических пингвинов к 10 годам
жизни на двух самцов остается только одна самка.
У некоторых летучих мышей после зимовки доля
самок может снижаться до 20%. У других видов,
наоборот, более высока гибель самцов (ряд
грызунов, утки-кряквы, большая синица и др.).
Половая структура популяции, естественно,
отражается на возможностях и темпах размножения
и роста численности.
Возрастная структура популяций также
тесно связана с динамикой их численности.
Биологические особенности видов определяют
основные три типа такой структуры.
У видов с короткой продолжительностью
жизни, высокой плодовитостью и однократным
размножением (однолетние растения, многие
насекомые, некоторые проходные рыбы) популяции,
как правило, одновозрастные. Очередное
поколение практически одновременно появляется
из яиц или семян, более или менее синхронно
подрастает, переходит к размножению, после
которого отмирает. У таких видов материнское
поколение не встречается с дочерним. Например, у
всех известных саранчовых перезимовывают яйца,
отложенные в почву в «кубышках», а взрослые
насекомые к зиме погибают. Так как нежные, только
что вышедшие из яиц насекомые наиболее
чувствительны к засухе, заморозкам и недостатку
корма, то они в массе гибнут при неблагоприятных
погодных условиях. Суровые зимы также
сказываются на выживании яиц. Поэтому
численность популяций саранчовых подвержена
резким изменениям в разные годы. Точно так же
посевы однолетних растений наиболее уязвимы на
стадии всходов. Динамика численности всех видов
с упрощенной возрастной структурой популяции
отличается резкими флуктуациями и
нестабильностью. Периодически такие популяции
могут давать очень резкие вспышки численности.
Другой тип возрастной структуры
популяций – прямо противоположный: присутствуют
все возрастные группы, одновременно существует
несколько поколений. Члены таких популяций в
большей степени отличаются друг от друга по
устойчивости к неблагоприятным факторам,
действие которых, как правило, приводит к потерям
только некоторых особей. Такие популяции
характерны для видов с большой
продолжительностью жизни и неоднократным
размножением (слоны, лошади, большинство птиц,
многолетние травы, деревья и т.п.). К этим видам
относится и человек. Динамика численности
подобных популяций обычно сглажена, без резких
колебаний.
Промежуточный характер имеют
популяции видов с небольшой продолжительностью
жизни, но повторным размножением (например, у
мышевидных грызунов). У них часть сезона
популяция почти одновозрастна, а с началом
размножения быстро пополняется особями разных
возрастных групп с появлением и дочерних, и
внучатых особей. Однако к концу сезона
размножение прекращается, и перезимовывают лишь
зверьки, родившиеся осенью или в конце лета. Для
таких популяций также характерна неустойчивая
численность и ее периодические подъемы и спады.
Возрастная структура популяции любого
вида подвержена влиянию внешних условий (погоды,
врагов и т.п.), что отражается на возможностях
размножения и пополнения популяции в ближайшем
будущем. Это обстоятельство широко используется
для прогноза численности интересующих нас видов,
а также человеческого населения. В частности,
если в разновозрастной популяции соотношение
молодых, взрослых и старых особей отразить в виде
расположенных друг над другом прямоугольников,
то для растущей популяции они образуют пирамиду
с широким основанием и узкой вершиной. В этом
случае число молодых особей значительно, а число
старых особей – меньше, чем зрелых, и постепенно
убывает. При примерном равенстве числа молодых и
размножающихся взрослых пирамида отражает
стабильную популяцию. Если же ее основание более
узкое, чем середина или вершина, популяции грозит
в ближайшее время сокращение общей численности.
Такие демографические пирамиды, построенные с
разной степенью дробности учета возраста, широко
используются в моделях, прогнозирующих
структуру населения государств, регионов и
человечества в целом.
Анализ возрастной структуры популяции
очень важен для грамотного использования
промысловых видов и для защиты редких и
исчезающих видов. В рыболовстве для видов с
неустойчивой численностью характерны
«урожайные» и «неурожайные» годы. У трески,
сельди, например, численность мощных поколений
может быть в 50–90 раз больше слабых, поэтому очень
важно устанавливать и соблюдать квоты на размеры
вылавливаемых рыб, чтобы не подорвать
возможности восстановления стада (вылавливая в
основном «урожайные» поколения).
Многолетние растения проходят разные
стадии своего развития – от проростков до
генеративных и затем стареющих особей – с
неодинаковой скоростью. У ряда видов в
популяциях нормально преобладают молодые, у
других – зрелые и даже стареющие экземпляры со
слабой способностью к семенному
воспроизводству. Это зависит от того, сколько
времени данный вид растения задерживается на
генеративной стадии и как быстро он ее достигает.
Например, у ковыля от прорастания семени до
полного отмирания всех вегетативных частей
проходит несколько десятилетий, иногда – до 100 и
более лет. Большую часть этого времени растения
проводят в состоянии мощной дерновины, ежегодно
возобновляя вегетацию и давая семена, а время от
прорастания до первого цветения занимает лишь
неcколько лет. Нормальный возрастной спектр
популяции в данном случае содержит малую долю
молодых и большую – зрелых и старых растений.
Многочисленным семенам негде прорастать, т.к.
место занято родительскими особями, они
резервируются в почве, образуя надежный запас, и
прорастают при любом освобождении напочвенного
покрова, пополняя популяцию. Отклонение от
обычного возрастного состава, которое
называется базовым возрастным спектром,
всегда связано с влиянием экологических
факторов. Например, при усилении выпаса может
происходить омоложение популяции многолетних
злаков, т.к. скот разрывает копытами дерновину,
освобождая место для прорастающих семян. Но
избыток молодых растений делает популяцию
неустойчивой, и ее судьба зависит от того, будет
ли выпас ослаблен или временно прекращен.
Территория, занимаемая популяцией,
содержит ресурсы для жизни. Количество ресурсов,
способных поддержать максимально возможную в
данных условиях плотность популяции, носит
название емкость среды. Полнота
использования ресурсов зависит не только от
численности, но и от размещения особей в
пространстве. В ходе эволюции у видов
выработались разные способы освоения
территории, позволяющие поддерживать на ней
определенную численность, не создавая угрозы
перенаселения. Пространственная структура
популяции создается за счет взаимодействия
членов популяции, их реакции друг на друга при
освоении среды.
Растения занимают среду, пронизывая
почву корневой системой и располагая
фотосинтезирующие органы на определенной высоте
в соответствующем объеме воздуха. Площадь их
питания не может быть меньше некоторой
определенной величины. В ходе роста они остро
конкурируют друг с другом, и в результате на
единице площади остается то количество
экземпляров, которое соответствует емкости
среды. (Гектар елового леса содержит 600–700
взрослых деревьев, хотя всходы молодых елочек
могут исчисляться десятками тысяч.)
Подвижность животных позволяет им
иными способами рационально использовать
территорию, сочетая элементы конкурентных и
взаимовыгодных отношений. У них различают
оседлый и кочевой образ жизни. Оседлые животные
характеризуются привязанностью к определенному
участку местообитания. Эта территориальная
привязанность носит название хоминг, т.е.
чувство дома. Для оседлых животных характерно
территориальное поведение, которое включает две
составляющие: охрану занимаемого участка и его
активное исследование и использование. Участок
охраняется от соседей, и таким образом
занимаемая популяцией территория оказывается
поделенной на отдельные ячейки, достаточные для
того, чтобы прокормить отдельную особь или
семейную группу. Охрана индивидуальных участков
осуществляется по-разному. В крайних проявлениях
– это прямые схватки с претендентами, но чаще –
ритуальные угрозы, звуковые сигналы или пахучие
и визуальные метки, свидетельствующие, что
участок занят. В инстинктах животных заложено
избегание занятых участков, на чужой территории
у них преобладает ориентировочное поведение, а
на своей – агрессивное.
Использование ресурсов у
территориальных животных носит интенсивный
характер. Досконально изучив свой участок,
хозяин знает, где и в какое время искать корм,
куда спрятаться в случае опасности, как
защититься от непогоды, строит гнезда, логова,
убежища или систему нор. Оседлость может быть
постоянной (сурки, суслики, пищухи и др.) или
приуроченной только к периоду размножения
(многие птицы, некоторые рыбы и др.).
В территориальном поведении
присутствуют элементы конкуренции. Однако
соседи связаны и мутуалистическими отношениями.
Они проявляются в оповещении друг друга о
замеченной опасности, в объединении против
общего врага. В популяции постоянен поток
информации о местопребывании отдельных особей, о
занятости территории. К ним добавляются сигналы
тревоги, реакции испуга, половые призывы.
Стереотипы поведения животных направлены на
поиск и поддержание контактов с себе подобными.
Если численность популяции низка и вокруг много
свободного пространства, животные не селятся
поодиночке, а выбирают участки, расположенные по
соседству. Та сумма информации, которую они
получают друг о друге, получила название биологическое
сигнальное поле.
Максимальный размер участка
определяется не только его кормностью, но и
возможностями ориентироваться на поведение
соседей. На границах индивидуальных участков,
например, у сусликов, часто имеются нейтральные
зоны, где соседи общаются друг с другом без
агрессии, а молодые могут играть друг с другом.
Площадь индивидуальной территории
может меняться в зависимости от плотности
популяции и урожайности года, но существуют
минимальные пределы, необходимые для выживания
хозяев. Те особи, которым не достались участки,
расселяются на другие, менее удобные, территории
или не приступают к размножению.
Альтернативный тип использования
пространства – кочевой образ жизни. Животные
передвигаются по достаточно обширной
территории, меняя места обитания и собирая
пищевые ресурсы. Это экстенсивный способ
освоения среды. Он снимает зависимость от
недостатка кормов на отдельных участках. Однако
такой образ жизни усиливает опасность гибели от
врагов или других неожиданных обстоятельств и
оказывается не под силу одиночным особям.
Животные кочуют только группами (слоны, копытные,
дельфины, павианы и др.). В группах
устанавливаются определенные отношения,
выявляются лидеры или вожаки, которым подражают
или подчиняются другие особи, осуществляется
защита детенышей. Группа использует опыт и силу
наиболее полноценных своих членов и действует
как целое. В стаи без вожаков животных (рыб,
насекомых) объединяет подражательное поведение
и система информационных сигналов. Отношения
животных в группах (стадах, стаях, колониях) имеют
характер социальных связей, они также
представляют собой переплетение конкуренции и
мутуализма.
Животные кочуют в пределах
определенных территорий, возвращаясь на прежние
места по мере восстановления там кормовых
запасов. Для зебр в саваннах Африки кочевые
территории в среднем составляют пространство в
400 км2 (20х20 км), а для северных оленей маршруты
кочевок могут быть протяженностью до 1000 км, от
тундр на побережье Ледовитого океана до полосы
лесотундры или северной тайги. Маршруты эти
постоянны и в настоящее время часто нарушаются
строительством промышленных сооружений (нефте- и
газопроводов, автомобильных трасс и т.п.).
Отклонения от привычных маршрутов приводят к
недокорму и голодовкам животных, нанося
существенный урон промысловому оленеводству.
У животных, проводящих всю жизнь в
стадах, стаях, колониях или скоплениях,
существует выраженный эффект группы.
Физиологическое состояние отдельного животного
резко ухудшается, если оно вынуждено
существовать в одиночку или с малым числом
сородичей. Колониальные птицы, если, например, их
число невелико, не приступают к размножению
после разорения птичьего базара. Рыбы стайных
видов (например, сельди) не питаются даже при
обилии корма, если вокруг нет сородичей. У
муравьев, пчел, отселенных от общины, резко
сокращается продолжительность жизни и угасают
некоторые важные инстинкты даже в оптимальных, с
физической точки зрения, условиях.
Необходимость существования в группе
сильно осложняет возможности восстановления
вымирающих видов. С ней необходимо считаться и в
животноводстве, и при охране промысловых
животных. Этот же эффект приходится учитывать и
при борьбе с вредителями. Например, если кладки
яиц сибирского шелкопряда сохраняются почти
полностью из-за низкой активности естественных
врагов, гусеницы развиваются группами вдвое
быстрее, чем поодиночке, и гораздо более
прожорливы.
Расселение – одна из важнейших
функций любого вида. Попадая на новые, еще не
освоенные территории, небольшое количество
особей, в случае успешного размножения,
основывают новые популяции.
Рост популяций у любых видов, от
бактерий до человека, имеет общие
закономерности. Теоретически численность любого
вида может расти бесконечно в силу того, что
размножение обычно происходит в геометрической
прогрессии. Число потомков, которое может
оставить после себя одна особь, носит название биотический
потенциал вида. Потомков может быть всего
два, как при простом делении клетки надвое
(бактерии, амебы, инфузории), а может быть – сотни
миллионов (количество икринок, откладываемых за
жизнь рядом плодовитых рыб, число мелких семян у
многих древесных растений и т.п.). Но даже при
простом удвоении за поколение кривая
численности популяции носит характер
экспоненты, т.е. быстро взмывает вверх, в
бесконечность. У амеб или бактерий
геометрическая прогрессия размножения может
дать уже через 20 делений из одной клетки – более
миллиона особей, и по подсчетам превысить массу
земной коры всего за полтора-двое суток. Такой
мощный потенциал размножения, которым обладают
живые существа, никогда не реализуется полностью
прежде всего из-за нехватки ресурсов, которые для
любого вида на планете имеют пределы, а в
конкретных местообитаниях соответствуют
емкости среды.
Кривая численности популяции в новых
для нее условиях сначала поднимается вверх, а
затем тормозится на определенном уровне и
колеблется около него, отражая влияние различных
факторов. В математике рост популяции отражается
так называемыми логистическими уравнениями,
показывающими, что с увеличением численности
популяции скорость прироста замедляется,
приближаясь к нулю, и кривая поворачивает
параллельно горизонтальной оси. Так растут
культуры бактерий в пробирках, завезенные на
поле сорняки, стада разводимых
сельскохозяйственных животных, население вновь
осваиваемых районов и в целом все человечество
на планете.
Сдерживание роста популяций в природе
происходит в результате действия целого ряда
факторов. Их делят на две принципиально
различные категории: модификаторы и регуляторы. Факторы-модификаторы
воздействуют на численность популяции
односторонне, сами не испытывая никакого влияния
с ее стороны. Это прежде всего – абиотические
явления: засухи, проливные дожди, наводнения,
бури, низкие температуры и т.п. Все знают,
насколько зависят урожаи культурных растений от
таких явлений и как экстремальные ситуации
усложняют выживаемость не только диких животных,
но и людей. Метеорологические прогнозы очень
важны и для сельского хозяйства, и для медицины.
Рост и плодовитость особей подчиняется правилу
оптимума, поэтому все отклонения от него
изменяют (модифицируют) численность популяций в
сторону уменьшения, а приближение к оптимуму
стимулирует увеличение численности.
Факторы-регуляторы не только
влияют на численность популяций, но и сами
изменяют силу своего действия в зависимости от
ее плотности. С падением численности воздействие
регуляторов ослабевает, с возрастанием –
усиливается. Возникает так называемая отрицательная
обратная связь, которая способна сдерживать
рост популяции. Следует отметить, что чрезмерный,
неконтролируемый, рост численности в конечном
итоге губителен для любого вида, т.к. может
полностью истощить и подорвать ресурсы среды. В
эволюции возникло множество механизмов
регулирования размножаемости видов.
Факторы регуляторы в природе делятся
на две группы: биоценотические и внутривидовые
(популяционные). Биоценотические регуляторы
численности отдельных популяций – это враги:
хищники, паразиты, возбудители болезней, а также
конкуренты из числа других видов. С увеличением
плотности популяций сила их действия возрастает,
а при низкой плотности – ослабевает. Хищники
переключаются на более доступную добычу, а
распространение паразитов от одного хозяина к
другому затрудняется. Пресс врагов и конкурентов
– мощные силы сдерживания роста популяций.
Например, при плодовитости взрослой трески до 10
млн икринок за один нерест до взрослого
состояния в лучшем случае доживают одна или
несколько особей. Меньшая часть икринок и
мальков гибнет под влиянием абиотических
факторов, а бoльшая – поедается разного рода
фильтраторами, мелкими и крупными хищниками.
Паразиты присутствуют в популяциях рыб в большом
количестве и разнообразии, снижая их
плодовитость и выживаемость. Среди врагов
сибирского шелкопряда, способного оголять
хвойные леса, наиболее эффективные
биоценотические регуляторы – мелкие
перепончатокрылые насекомые, паразиты-яйцееды.
Они уничтожают до 95% яиц в кладках шелкопряда.
Гусеницы и взрослые бабочки также испытывают
пресс хищников. В период активности регуляторов
вспышек численности шелкопряда не происходит.
Наши предки-собиратели также страдали
от хищников, в основном, от крупных кошачьих:
леопардов, львов и др. Для современного человека
этот фактор практически полностью устранен, а
успешную борьбу с паразитами и инфекционными
заболеваниями ведет медицина.
Внутривидовые (популяционные)
механизмы регуляции имеются практически у
всех видов, они действуют наряду с
биоценотическими, межвидовыми и выступают на
первый план у тех видов, у которых в природе мало
естественных врагов. Они поддерживают гомеостаз
– примерное постоянство численности популяций,
препятствуя ее (численности) чрезмерному
увеличению. Разные по экологии и биологии виды
сильно различаются по способам саморегуляции.
У растений, не способных менять свое
местопребывание, возникает острая конкуренция
за площадь питания и свет, в результате сильные
особи губят слабых, перехватывая средства
существования. Происходит самоизреживание,
которое в конечном счете идет на пользу виду,
поскольку сохраняются генетически и
физиологически более полноценные особи.
Другой способ торможения роста
численности – отравление среды продуктами
обмена веществ, которые ингибируют (подавляют)
размножение. Он развивается у видов, населяющих
относительно замкнутое пространство, где
возможно накопление таких веществ: в культурах
бактерий, у рыб и земноводных в непроточных
водоемах, у насекомых – вредителей запасов зерна
и т.п. В биотехнологиях, основанных на
выращивании одноклеточных и мелких организмов в
водных растворах, их продукция резко повышается
при создании проточных условий. В аквариумах
рыбы не растут, если их поместить в воду, где до
этого было много рыб того же вида.
Наиболее распространенный и
эффективный способ поддержания популяционного
гомеостаза у животных – их территориальное
поведение. Этот способ регуляции плотности
населения имеется в самых разных группах: у
млекопитающих, птиц, рептилий, ряда рыб, у пауков,
многих насекомых (особенно с общественным,
социальным, образом жизни – муравьев, пчел,
термитов). Защита и охрана индивидуальных и
семейных участков не позволяет возникнуть
избытку особей, угрожающему подрывом общих
ресурсов.
Миграции и расселение животных –
также распространенный способ снятия угрозы
перенаселенности. Инстинкты миграции
усиливаются при увеличении плотности популяции.
Наиболее яркий пример – норвежские лемминги,
которые в годы вспышек численности сплошным
потоком начинают переселяться из мест рождения,
преодолевая различные преграды и в массе
погибая. Еще более впечатляющи миграции стадной
саранчи на сотни и тысячи километров. Насекомые
образуют тучи в тысячи тонн суммарного веса,
поедают на своем пути всю растительность, губят
урожаи, но и сами обречены на гибель, т.к. в местах
оседания, как правило, не находят условий для
размножения. Образование перелетных стай – это
реакция саранчи на увеличение плотности своего
вида в местах постоянного обитания. При этом
меняется физиологическое и гормональное
состояние особей, характер поведения,
проявляющиеся инстинкты и даже внешний облик.
Насекомые переходят через ряд линек из так
называемой одиночной фазы в стадную, собираются
вместе и улетают, разреживая оставшуюся часть
популяции.
Повышение плотности популяции
изменяет поведение, снижает плодовитость и
увеличивает смертность у многих групп животных.
У млекопитающих при переуплотнении населения
возникает стресс-реакция – нервно-гормональные
изменения в организме, блокирующие размножение.
Эти изменения обратимы, при уменьшении
численности состояние животных вновь
нормализуется.
Таким образом, внутрипопуляционные
способы регуляции численности, сформированные
естественным отбором, очень разнообразны у
различных видов, но приводят к сходным
результатам – предотвращению полного подрыва
ресурсов и сохранению вида на данной территории.
Они представляют собой реакцию на повышающуюся
плотность населения и проявляются до того, как
произойдет исчерпание емкости среды.
Как биоценотические, так и
популяционные механизмы регуляции при повышении
плотности популяции могут включаться быстро, но
могут и запаздывать, проявляя инерцию. Например,
размножение хищников вслед за размножением
жертв – всегда запаздывающий процесс. Изменение
плодовитости также сказывается на численности
популяции не сразу, а только в следующем
поколении. Поэтому, несмотря на многочисленные
регуляторные связи, популяции на некоторый
период могут выходить из-под действия
регуляторных механизмов – тогда происходят
вспышки численности. На эту динамику
накладывается также влияние односторонне
действующих, модифицирующих факторов. Поэтому
численность популяций всегда варьирует, более
или менее отклоняясь от средней.
Различают три основных типа динамики
численности популяций: стабильный,
флуктуирующий и взрывной. В первом случае виды
находятся под влиянием эффективных регуляторов,
и колебания численности не превышают нескольких
крат. Во втором случае размах колебаний выше – на
порядок величин, но преобладают безынерционные
регуляторные механизмы, быстро снижающие
повышенную плотность населения. Взрывной ход
численности характерен для видов, у которых
регуляторные механизмы обладают запаздывающим
действием. Размах колебаний численности может
при этом достигать двух, а иногда и трех порядков
величин (лемминги, сибирский и непарный
шелкопряды, луговой мотылек и др.).
Антропогенное влияние на динамику
численности видов чаще всего выражается в
нарушении регуляторных механизмов. Изменяя
биотопы и обедняя сообщества, человек, чаще всего
неосознанно, снимает заслоны, стоящие на пути
размножения многих видов. Если исчезают враги,
паразиты и конкуренты, нарушаются
территориальные связи, динамика вида из
стабильной может превратиться во взрывную.
Возникают проблемы вредителей леса, сельского
хозяйства и различных запасов, проблемы
распространения инфекций и т.п.
Демографические проблемы весьма
актуальны и для современного человечества.
Последние 200 лет характеризуются таким ростом
населения Земли, что он получил название демографический
взрыв. За это время численность людей
увеличилась в 6 раз и в настоящее время
приближается к
7 млрд человек. В эпоху собирательства, по всей
вероятности, численность человечества
составляла не более 50 млн, а к началу нашей эры не
превышала 300 млн. Увеличение емкости среды было
обусловлено развитием цивилизации. Основными
ограничителями плотности населения древних
людей были недостаток пищи и малая
продолжительность жизни, которая не охватывала
всего репродуктивного периода. Первое
значительное увеличение емкости среды для людей
произошло 8–12 тыс. лет назад в связи с переходом
от собирательства и охоты к производящему
хозяйству – выращиванию растений и
животноводству (неолитическая революция).
Следующий значительный подъем численности был
связан с промышленной революцией XVIII в., когда
труд людей был облегчен и интенсифицирован
использованием машинной техники и горючих
ископаемых. К концу этого века на Земле
насчитывалось уже 800 млн человек. После
научно-технической революции XIX в. успехи
медицины и улучшение условий жизни снизили
смертность, и средняя продолжительность жизни
впервые охватила весь репродуктивный период.
Рождаемость продолжала оставаться высокой,
темпы прироста населения возросли.
Об угрозе перенаселенности Земли
впервые заговорил английский пастор Томас
Мальтус в конце XVIII в., предрекая грядущие
катастрофы. Однако по-настоящему
обеспокоенность этой проблемой охватила
общественность в 60-е гг. XX в., когда стало ясно, что
численность людей растет угрожающими темпами и
мир приближается к исчерпанию ресурсов и
тотальному загрязнению. Возникла Международная
программа планирования семьи, пропагандирующая
сокращение рождаемости, которая достаточно
жестко проводилась в жизнь в наиболее населенных
странах – Китае и Индии. Компьютерные прогнозы
развития человечества, обнародованные Денисом и
Донеллой Медоузами в 1972 г. на Стокгольмской
международной конференции по состоянию
окружающей среды, предрекали всеобщую
экологическую катастрофу через считанные
десятилетия. Эта угроза продолжает оставаться
реальной и в наше время. Однако в темпах прироста
населения наметились некоторые изменения.
Численность человечества продолжает расти, но
уже несколько замедляется. Темпы прироста
достигали максимальных величин в 1970-е гг. – 2,06% за
год, в 2000 г. они составляли 1,35%, в 2005 – 1,22%. Кривая
демографического роста постепенно приближается
к логистической, что является закономерным
результатом для всех растущих популяций. По
расчетам, численность человечества может
стабилизироваться в XXII в. на уровне 10–11 млрд
человек. При этом изменится соотношение
численности народов мира. Демографические
процессы должны характеризоваться низкой
рождаемостью и низкой смертностью при
увеличении продолжительности жизни.
Природа Земли сможет прокормить такое
количество людей, но только при условии
рационального, экологически грамотного
хозяйствования, более справедливого социального
распределения пищевых и прочих ресурсов и
повышения образованности населения. За
осуществление этих условий предстоит сложная и
напряженная социальная борьба и в отдельных
странах, и в целом мировом сообществе.
Вопросы и задания для самостоятельной работы
1. Что такое популяции? Каковы их
основные характеристики?
2. Каковы теоретические и реальные
особенности роста популяций?
3. Какие типы возрастной и половой
структуры популяций характерны для разных видов?
4. Какие пути предотвращения
перенаселенности (механизмы гомеостаза
популяций) возникли в эволюции разных видов?
Какие еще причины сдерживают рост популяций?
5. Что такое «демографический взрыв» в истории
человечества? Почему он происходит и каковы
связанные с ним проблемы?
Литература
1. Шилов И.А. Экология. Учебник для
биол. и мед. вузов. – М.: Высшая школа, 1997–2003.
2. Чернова Н.М., Былова А.М. Общая
экология. Учебник для студентов педагогических
вузов. – М.: Дрофа, 2004.
3. Гиляров А.М. Популяционная
экология. – М.: изд-во МГУ, 1990.
Для описания популяции используются статистические динамические характеристики, присущие всей совокупности особей, относящихся к описываемому виду. К основным статистическим характеристикам популяции относятся следующие: численность популяции, плотность популяции, показатели структуры.
Под численностью популяции понимают количество особей растений и животных, принадлежащих одному виду, которые включены в состав популяции и населяют определенную территорию в биогеоценозе).
Численность популяций зависит от размера особей и величины популяционного ареала. Численность популяции характеризуется: кривыми выживания, кривыми роста, кривыми численности. Методы определения численности: подсчет особей при помощи аэрофотосъемки, метод мечения, метод кольцевания.
Плотность популяции — это величина, измеряемая числом особей или биомассой популяции на единицу площади или объема территории обитания.
Различают среднюю плотность (число особей на единицу всего пространства) и экологическую плотность (число особей на единицу обитаемого пространства).
Плотность популяции изменяется в зависимости от численности. При возрастании численности плотность популяции не увеличивается лишь в том случае, если возможно расселение особей и расширение ее популяционного ареала. Максимальная плотность для различных видов организмов и условий существования сильно отличается. Показатель плотности популяции позволяет количественно сравнивать отдельные популяций, независимо от общего размера занимаемой ими территории или акватории. Кроме того, зная плотность популяции в тот или иной момент времени, можно судить о росте, размножении и старении популяции.