Решение «популярных» задач по физике на тему «Потенциал»
Разберем сегодня несколько задач по физике, связанных с вычислением потенциала сферы. Так уж сложилось, что новые задачи по физике появляются значительно реже, нежели, к примеру скажем, по математике. Оно и понятно, ведь придумать оригинальную физическую задачу далеко не просто. Из года в год в различных физических олимпиадах, вариантах ЕГЭ по физике и других диагностических работах появляются одни и те же задачи, причем зачастую авторы по разным причинам не меняют даже числовых значений входящих в условие параметров. Решение некоторых таких часто встречающихся (напрашивается назвать их «бородатыми», но мы назовем их лучше «популярными») задач приведено в данной статье.
Задача 1. В одну большую каплю сливают n одинаковых капелек ртути, заряженных до потенциала φ. Каков будет потенциал Φ этой капли? Считать, что капли имеют сферическую форму.
Решение. Потенциал заряженного шара (коим по условию является каждая из капель) определяется по формуле:
![[ varphi = frac{Q}{4pivarepsilon_0 R}, ]](https://yourtutor.info/wp-content/ql-cache/quicklatex.com-763b3caa8fc94a645e7a4b1f728c7849_l3.png)
где Q — заряд шара, ε0 = 8,85 · 10 -12 Ф/м — диэлектрическая постоянная, R — радиус шара.
Тогда потенциал образовавшейся после слияния капли можно определить так:
![[ Phi = frac{Q}{4pivarepsilon_0 R}. ]](https://yourtutor.info/wp-content/ql-cache/quicklatex.com-2db31d21e9e91a76459e0d6192318562_l3.png)
Общий заряд Q, по закону сохранения заряда, определяется суммой зарядов q каждой маленькой капли: Q = n · q. Как связать радиус R получившейся большой капли с радиусом r каждой малой? Используем то, что в результате слияния не изменяется объем ртути, то есть (предполагается, что вы помните формулу для вычисления объема шара, если нет — загляните сюда):
![[ ncdotfrac{4}{3}pi r^3 = frac{4}{3}pi R^3Leftrightarrow R = rsqrt[3]{n}. ]](https://yourtutor.info/wp-content/ql-cache/quicklatex.com-278d0c50065aff2b973634421180db2d_l3.png)
![[ Phi = frac{nq}{4pivarepsilon_0 rsqrt[3]{n}} = frac{qsqrt[3]{n^2}}{4pivarepsilon_0 r}. ]](https://yourtutor.info/wp-content/ql-cache/quicklatex.com-6822970ce3b2b54dd87cdc0777bfda52_l3.png)
![[ varphi = frac{q}{4pivarepsilon_0 r} ]](https://yourtutor.info/wp-content/ql-cache/quicklatex.com-e77c5408b73f639888c8252b18ba5a65_l3.png)
есть по определению потенциал одной маленькой капли, поэтому окончательно получаем ответ:
![[ Phi = varphisqrt[3]{n^2}. ]](https://yourtutor.info/wp-content/ql-cache/quicklatex.com-b1c92ff058a80c5f60f9c362d58d3930_l3.png)
Задача 2. Металлический шар радиусом r помещен в жидкий диэлектрик с плотностью ρ2. Плотность материала, из которого изготовлен шар, ρ1 (ρ1 > ρ2). Чему равен заряд шара, если в однородном электрическом поле, направленном вертикально вверх, шар оказался взвешенным в жидкости? Электрическое поле создается двумя параллельными пластинами, расстояние между которыми d, а разность потенциалов U.
Решение.
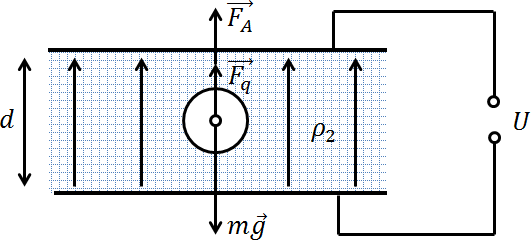
Поскольку шар находится в равновесии, векторная сумма всех сил, действующих на него, равна нулю
На шар действуют три силы: сила тяжести mg = ρ1gV (направлена вниз), выталкивающая сила Архимеда FA = ρ2gV (направлена вверх), сила Кулона Fq = qE (направлена вверх). То, что сила Кулона направлена вверх следует из того, что плотность материала шара больше плотности жидкого диэлектрика, в котором он плавает. Это означает, что он утонул бы, не будь он заряжен. Спасает его от этого дополнительная сила Кулона, сонаправленная с выталкивающей силой Архимеда.
Шар находится в равновесии, значит векторная сумма всех действующих на него сил равна нулю:
![[ mvec{g}+vec{F_q}+vec{F_A}. ]](https://yourtutor.info/wp-content/ql-cache/quicklatex.com-31c6a9fd6a1c7f45e2efa181f2296c28_l3.png)
Или в проекции на вертикальную ось:
![[ mg = F_A+F_q. ]](https://yourtutor.info/wp-content/ql-cache/quicklatex.com-624b7ccf234b59bd556897d9ecfdd32c_l3.png)
С учетом написанных выше формул:
![[ rho_1 gV = rho_2 gV + qELeftrightarrow q=frac{(rho_1-rho_2)gV}{E}. ]](https://yourtutor.info/wp-content/ql-cache/quicklatex.com-54d18b3bc80ad10307f64af57f4e8f29_l3.png)
С учетом формулы для объема шара (V = 4/3πr 3 ) и формулы, отражающей взаимосвязь напряженности поля с напряжением между двумя точками (U=E·d), получаем окончательный ответ:
![[ q = frac{4pi r^3gd(rho_1-rho_2)}{3U}. ]](https://yourtutor.info/wp-content/ql-cache/quicklatex.com-82376bebf6041ba3a526c194fe06bd3c_l3.png)
Задача 3. Проводник длиной l движется с постоянным ускорением a, направленным вдоль его оси. Определите напряжение, возникающее между концами проводника; me — масса электрона, |e| — элементарный заряд.
Решение. В процессе движения стержня часть электронов по инерции смещается к одному из его концов (ситуация напоминает поезд в метро — стержень — и едущих в нем пассажиров — электроны).
Процесс перетекания будет продолжаться до тех пор, пока возникшее в стержне электрическое поле не начнет действовать на электроны с силой |e|E, где E — напряженность этого поля, равной по величине mea. Напряженность поля связана с напряжением между концами проводника соотношением: U = E · l. После всех подстановок и преобразований получает ответ:
![[ U = frac{m_eal}{|e|}. ]](https://yourtutor.info/wp-content/ql-cache/quicklatex.com-99f8644e3fa45ffb094a3cac42d3d5bf_l3.png)
Задачи взяты из сборника В. И. Васюкова, С. Н. Дмитриева, Ю. А. Струкова «Физика: сборник задач для поступающих в вузы». Все задания в этом сборнике приведены с ответами, поэтому при желании вы можете самостоятельно оценить свои силы в их решении. Присылайте свои вопросы и интересные задачи, и мы обязательно разберем их в одной из следующих статей.
Формирование мембранного потенциала покоя
 Обзор
Обзор
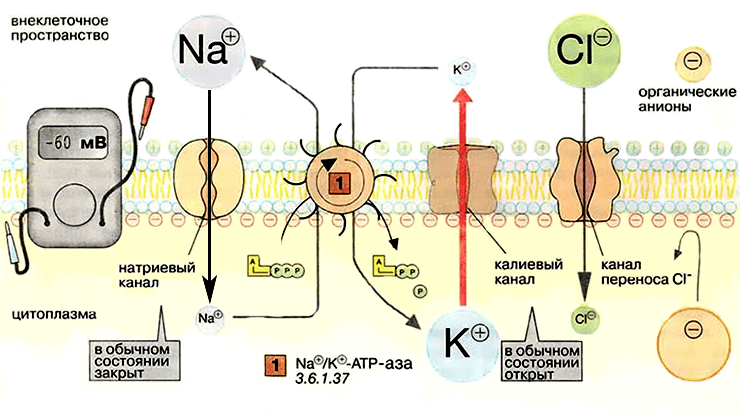
Потенциал покоя. В мембране открыты калиевые ионные каналы постоянного тока, закрыты натриевые каналы, работает насос-обменник (Na + /K + -АТФаза).
Автор
Редакторы
- Ионные каналы
- Нейробиология
Статья на конкурс «био/мол/текст»: Потенциал покоя — это важное явление в жизни всех клеток организма, и важно знать, как он формируется. Однако это сложный динамический процесс, трудный для восприятия целиком, особенно для студентов младших курсов (биологических, медицинских и психологических специальностей) и неподготовленных читателей. Впрочем, при рассмотрении по пунктам, вполне возможно понять его основные детали и этапы. В работе вводится понятие потенциала покоя и выделяются основные этапы его формирования с использованием образных метафор, помогающих понять и запомнить молекулярные механизмы формирования потенциала покоя.

«Био/мол/текст»-2011
Эта статья представлена на конкурс научно-популярных работ «био/мол/текст»-2011 в номинации «Лучшая обзорная статья».
Мембранные транспортные структуры — натрий-калиевые насосы — создают предпосылки для возникновения потенциала покоя. Предпосылки эти — разность в концентрации ионов на внутренней и наружной сторонах клеточной мембраны. Отдельно проявляет себя разность концентрации по натрию и разность концентрации по калию. Попытка ионов калия (K + ) выровнять свою концентрацию по обе стороны мембраны приводит к его утечке из клетки и потере вместе с ними положительных электрических зарядов, за счёт чего значительно усиливается общий отрицательный заряд внутренней поверхности клетки. Эта «калиевая» отрицательность составляет бóльшую часть потенциала покоя (−60 мВ в среднем), а меньшую его часть (−10 мВ) составляет «обменная» отрицательность, вызванная электрогенностью самого ионного насоса-обменника.
Давайте разбираться подробнее.
Зачем нам нужно знать, что такое потенциал покоя и как он возникает?
Вы знаете, что такое «животное электричество»? Откуда в организме берутся «биотоки»? Как живая клетка, находящаяся в водной среде, может превратиться в «электрическую батарейку» и почему она моментально не разряжается?
На эти вопросы можно ответить только в том случае, если узнать, как клетка создаёт себе разность электрических потенциалов (потенциал покоя) на мембране.
Совершенно очевидно, что для понимания того, как работает нервная система, необходимо вначале разобраться, как работает её отдельная нервная клетка — нейрон. Главное, что лежит в основе работы нейрона — это перемещение электрических зарядов через его мембрану и появление вследствие этого на мембране электрических потенциалов. Можно сказать, что нейрон, готовясь к своей нервной работе, вначале запасает энергию в электрической форме, а затем использует ее в процессе проведения и передачи нервного возбуждения.
Таким образом, наш самый первый шаг к изучению работы нервной системы — это понять, каким образом появляется электрический потенциал на мембране нервных клеток. Этим мы и займёмся, и назовём этот процесс формированием потенциала покоя.
Определение понятия «потенциал покоя»
В норме, когда нервная клетка находится в физиологическом покое и готова к работе, у неё уже произошло перераспределение электрических зарядов между внутренней и наружной сторонами мембраны. За счёт этого возникло электрическое поле, и на мембране появился электрический потенциал — мембранный потенциал покоя.
Как известно из физики, электрические заряды (движущиеся и неподвижные) формируют в пространстве электромагнитное поле, которое влияет на тела, обладающие электрическим зарядом. С точки зрения электромагнетизма клеточную мембрану можно представить как плоский конденсатор, заполненный однородным диэлектриком из неполярных молекул. Если конденсатор заряжен, то внутри него возникает электрическое поле, обусловленное поверхностной плотностью заряда. На поверхности мембраны возникают некомпенсированные заряды: положительные у «отрицательной» поверхности и отрицательные — у «положительной» [6].
Таким образом, мембрана оказывается поляризованной. Это означает, что она имеет разный электрический потенциал наружной и внутренней поверхностей. Разность между этими потенциалами вполне возможно зарегистрировать.
В этом можно убедиться, если ввести внутрь клетки микроэлектрод, соединённый с регистрирующей установкой. Как только электрод попадает внутрь клетки, он мгновенно приобретает некоторый постоянный электроотрицательный потенциал по отношению к электроду, расположенному в окружающей клетку жидкости. Величина внутриклеточного электрического потенциала у нервных клеток и волокон, например, гигантских нервных волокон кальмара, в покое составляет около −70 мВ. Эту величину называют мембранным потенциалом покоя (МПП). Во всех точках аксоплазмы этот потенциал практически одинаков.
Ноздрачёв А.Д. и др. Начала физиологии [5].
Ещё немного физики. Макроскопические физические тела, как правило, электрически нейтральны, т.е. в них в равных количествах содержатся как положительные, так и отрицательные заряды. Зарядить тело можно, создав в нем избыток заряженных частиц одного вида, например, трением о другое тело, в котором при этом образуется избыток зарядов противоположного вида. Учитывая наличие элементарного заряда (e), полный электрический заряд любого тела можно представить как q = ±N×e, где N — целое число.
Потенциал электростатического поля φ определяется как отношение потенциальной энергии W пробного заряда q к величине этого заряда: φ = W/q, откуда следует, что потенциал численно равен потенциальной энергии, которой обладает в данной точке поля единичный положительный заряд. Единицей измерения потенциала служит вольт (1 В) [4]. В некоторых случаях потенциал электрического поля нагляднее определяется как физическая величина, численно равная работе внешних сил против сил электрического поля E при перемещении единичного положительного точечного заряда из бесконечности в данную точку. Последнее определение удобно записать следующим образом:
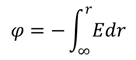
В электрофизиологии кроме потенциала покоя рассматриваются и другие электрические потенциалы: локальные постсинаптические и рецепторные потенциалы (возбуждающие и тормозные), электротонические и следовые потенциалы, миниатюрные потенциалы концевой пластинки, концентрационный потенциал и потенциал действия [5].
Потенциал покоя — это разность электрических потенциалов, имеющихся на внутренней и наружной сторонах мембраны, когда клетка находится в состоянии физиологического покоя. Его величина измеряется изнутри клетки, она отрицательна и составляет в среднем −70 мВ (милливольт), хотя в разных клетках может быть различной: от −35 мВ до −90 мВ.
Важно учитывать, что в нервной системе электрические заряды представлены не электронами, как в обычных металлических проводах, а ионами — химическими частицами, имеющими электрический заряд. И вообще в водных растворах в виде электрического тока перемещаются не электроны, а ионы. Поэтому все электрические токи в клетках и окружающей их среде — это ионные токи.
Итак, изнутри клетка в покое заряжена отрицательно, а снаружи — положительно. Это свойственно всем живым клеткам, за исключением, разве что, эритроцитов, которые, наоборот, заряжены отрицательно снаружи. Если говорить конкретнее, то получается, что снаружи вокруг клетки будут преобладать положительные ионы (катионы Na + и K + ), а внутри — отрицательные ионы (анионы органических кислот, не способные свободно перемещаться через мембрану, как Na + и K + ).
Теперь нам всего лишь осталось объяснить, каким же образом всё получилось именно так. Хотя, конечно, неприятно сознавать, что все наши клетки кроме эритроцитов только снаружи выглядят положительными, а внутри они — отрицательные.
Термин «отрицательность», который мы будем применять для характеристики электрического потенциала внутри клетки, пригодится нам для простоты объяснения изменений уровня потенциала покоя. В этом термине ценно то, что интуитивно понятно следующее: чем больше отрицательность внутри клетки — тем ниже в отрицательную сторону от нуля смещён потенциал, а чем меньше отрицательность — тем ближе отрицательный потенциал к нулю. Это намного проще понять, чем каждый раз разбираться в том, что же именно означает выражение «потенциал возрастает» — возрастание по абсолютному значению (или «по модулю») будет означать смещение потенциала покоя вниз от нуля, а просто «возрастание» — смещение потенциала вверх к нулю. Термин «отрицательность» не создаёт подобных проблем неоднозначности понимания.
Сущность формирования потенциала покоя
Попробуем разобраться, откуда берётся электрический заряд нервных клеток, хотя их никто не трёт, как это делают физики в своих опытах с электрическими зарядами.
Здесь исследователя и студента поджидает одна из логических ловушек: внутренняя отрицательность клетки возникает не из-за появления лишних отрицательных частиц (анионов), а, наоборот, из-за потери некоторого количества положительных частиц (катионов)!
Так куда же деваются из клетки положительно заряженные частицы? Напомню, что это покинувшие клетку и скопившиеся снаружи ионы натрия — Na + — и калия — K + .
Главный секрет появления отрицательности внутри клетки
Сразу откроем этот секрет и скажем, что клетка лишается части своих положительных частиц и заряжается отрицательно за счёт двух процессов:
- вначале она обменивает «свой» натрий на «чужой» калий (да-да, одни положительные ионы на другие, такие же положительные);
- потом из неё происходит утечка этих «наменянных» положительных ионов калия, вместе с которыми из клетки утекают положительные заряды.
Эти два процесса нам и надо объяснить.
Первый этап создания внутренней отрицательности: обмен Na + на K +
В мембране нервной клетки постоянно работают белковые насосы-обменники (аденозинтрифосфатазы, или Na + /K + -АТФазы), встроенные в мембрану. Они меняют «собственный» натрий клетки на наружный «чужой» калий.
Но ведь при обмене одного положительного заряда (Na + ) на другой такой же положительный заряд (K + ) никакого дефицита положительных зарядов в клетке возникать не может! Правильно. Но, тем не менее, из-за этого обмена в клетке остаётся очень мало ионов натрия, потому что они почти все ушли наружу. И в то же время клетка переполняется ионами калия, которые в неё накачали молекулярные насосы. Если бы мы могли попробовать на вкус цитоплазму клетки, мы бы заметили, что в результате работы насосов-обменников она превратилась из солёной в горько-солёно-кислую, потому что солёный вкус хлорида натрия сменился сложным вкусом довольно-таки концентрированного раствора хлорида калия. В клетке концентрация калия достигает 0,4 моль/л. Растворы хлорида калия в пределах 0,009–0,02 моль/л имеют сладкий вкус, 0,03–0,04 — горький, 0,05–0,1 — горько-солёный, а начиная с 0,2 и выше — сложный вкус, состоящий из солёного, горького и кислого [8].
Важно здесь то, что обмен натрия на калий — неравный. За каждые отданные клеткой три иона натрия она получает всего два иона калия. Это приводит к потере одного положительного заряда при каждом акте ионного обмена. Так что уже на этом этапе за счёт неравноценного обмена клетка теряет больше «плюсов», чем получает взамен. В электрическом выражении это составляет примерно −10 мВ отрицательности внутри клетки. (Но помните, что нам надо ещё найти объяснение для оставшихся −60 мВ!)
Чтобы легче было запомнить работу насосов-обменников, образно можно выразиться так: «Клетка любит калий!» Поэтому клетка и затаскивает калий к себе, несмотря на то, что его и так в ней полно. И поэтому она невыгодно обменивает его на натрий, отдавая 3 иона натрия за 2 иона калия. И поэтому она тратит на этот обмен энергию АТФ. И как тратит! До 70% всех энергозатрат нейрона может уходить на работу натрий-калиевых насосов. (Вот что делает любовь, пусть она даже и не настоящая!)
Кстати, интересно, что клетка не рождается с готовым потенциалом покоя. Ей его ещё надо создать. Например, при дифференцировке и слиянии миобластов потенциал их мембраны изменяется от −10 до −70 мВ, т.е. их мембрана становится более отрицательной — поляризуется в процессе дифференцировки. А в экспериментах на мультипотентных мезенхимальных стромальных клетках костного мозга человека искусственная деполяризация, противодействующая потенциалу покоя и уменьшающая отрицательность клеток, даже ингибировала (угнетала) дифференцировку клеток [1].
Образно говоря, можно выразиться так: Создавая потенциал покоя, клетка «заряжается любовью». Это любовь к двум вещам:
- любовь клетки к калию (поэтому клетка насильно затаскивает его к себе);
- любовь калия к свободе (поэтому калий покидает захватившую его клетку).
Механизм насыщения клетки калием мы уже объяснили (это работа насосов-обменников), а механизм ухода калия из клетки объясним ниже, когда перейдём к описанию второго этапа создания внутриклеточной отрицательности. Итак, результат деятельности мембранных ионных насосов-обменников на первом этапе формирования потенциала покоя таков:
- Дефицит натрия (Na + ) в клетке.
- Избыток калия (K + ) в клетке.
- Появление на мембране слабого электрического потенциала (−10 мВ).
Можно сказать так: на первом этапе ионные насосы мембраны создают разность концентраций ионов, или градиент (перепад) концентрации, между внутриклеточной и внеклеточной средой.
Второй этап создания отрицательности: утечка ионов K + из клетки
Итак, что начинается в клетке после того, как с ионами поработают её мембранные натрий-калиевые насосы-обменники?
Из-за образовавшегося дефицита натрия внутри клетки этот ион при каждом удобном случае норовит устремиться внутрь: растворённые вещества всегда стремятся выровнять свою концентрацию во всём объёме раствора. Но это у натрия получается плохо, поскольку ионные натриевые каналы обычно закрыты и открываются только при определённых условиях: под воздействием специальных веществ (трансмиттеров) или при уменьшении отрицательности в клетке (деполяризации мембраны).
В то же время в клетке имеется избыток ионов калия по сравнению с наружной средой — потому что насосы мембраны насильно накачали его в клетку. И он, тоже стремясь уравнять свою концентрацию внутри и снаружи, норовит, напротив, выйти из клетки. И это у него получается!
Ионы калия K + покидают клетку под действием химического градиента их концентрации по разные стороны мембраны (мембрана значительно более проницаема для K + , чем для Na + ) и уносят с собой положительные заряды. Из-за этого внутри клетки нарастает отрицательность.
Тут ещё важно понять то, что ионы натрия и калия как бы «не замечают» друг друга, они реагируют только «на самих себя». Т.е. натрий реагирует на концентрацию натрия же, но «не обращает внимания» на то, сколько вокруг калия. И наоборот, калий реагирует только на концентрацию калия и «не замечает» натрий. Получается, что для понимания поведения ионов надо по отдельности рассматривать концентрации ионов натрия и калия. Т.е. надо отдельно сравнить концентрацию по натрию внутри и снаружи клетки и отдельно — концентрацию по калию внутри и снаружи клетки, но не имеет смысла сравнивать натрий с калием, как это, бывает, делается в учебниках.
По закону выравнивания химических концентраций, который действует в растворах, натрий «хочет» снаружи войти в клетку; туда же его влечёт и электрическая сила (как мы помним, цитоплазма заряжена отрицательно). Хотеть-то он хочет, но не может, так как мембрана в обычном состоянии плохо его пропускает. Натриевые ионные каналы, имеющиеся в мембране, в норме закрыты. Если все же его заходит немножко, то клетка сразу же обменивает его на наружный калий с помощью своих натрий-калиевых насосов-обменников. Получается, что ионы натрия проходят через клетку как бы транзитом и не задерживаются в ней. Поэтому натрий в нейронах всегда в дефиците.
А вот калий как раз может легко выходить из клетки наружу! В клетке его полно, и она его удержать не может. Он выходит наружу через особые каналы в мембране — «калиевые каналы утечки», которые в норме открыты и выпускают калий [5, 7].
К + -каналы утечки постоянно открыты при нормальных значениях мембранного потенциала покоя и проявляют взрывы активности при сдвигах мембранного потенциала, которые длятся несколько минут и наблюдаются при всех значениях потенциала. Усиление К + -токов утечки ведёт к гиперполяризации мембраны, тогда как их подавление — к деполяризации. . Однако, существование канального механизма, ответственного за токи утечки, долгое время оставалось под вопросом. Только сейчас стало ясно, что калиевая утечка — это ток через специальные калиевые каналы.
Зефиров А.Л. и Ситдикова Г.Ф. Ионные каналы возбудимой клетки (структура, функция, патология) [7].
От химического — к электрическому
А теперь — ещё раз самое главное. Мы должны осознанно перейти от движения химических частиц к движению электрических зарядов.
Калий (K + ) положительно заряжен, и поэтому он, когда выходит из клетки, выносит из неё не только самого себя, но и положительный заряд. За ним изнутри клетки к мембране тянутся «минусы» — отрицательные заряды. Но они не могут просочиться через мембрану — в отличие от ионов калия — т.к. для них нет подходящих ионных каналов, и мембрана их не пропускает. Помните про оставшиеся необъяснёнными нами −60 мВ отрицательности? Это и есть та самая часть мембранного потенциала покоя, которую создаёт утечка ионов калия из клетки! И это — большая часть потенциала покоя.
Для этой составной части потенциала покоя есть даже специальное название — концентрационный потенциал [5]. Концентрационный потенциал — это часть потенциала покоя, созданная дефицитом положительных зарядов внутри клетки, образовавшимся за счёт утечки из неё положительных ионов калия.
Ну, а теперь немного физики, химии и математики для любителей точности.
Электрические силы связаны с химическими по уравнению Гольдмана. Его частным случаем является более простое уравнение Нернста, по формуле которого можно рассчитать трансмембранную диффузионную разность потенциалов на основе различной концентрации ионов одного вида по разные стороны мембраны. Так, зная концентрацию ионов калия снаружи и внутри клетки, можно рассчитать калиевый равновесный потенциал EK:
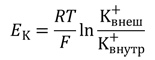
где Ек — равновесный потенциал, R — газовая постоянная, Т — абсолютная температура, F — постоянная Фарадея, К + внеш и K + внутр — концентрации ионов К + снаружи и внутри клетки, соответственно. По формуле видно, что для расчёта потенциала между собой сравниваются концентрации ионов одного вида — K + .
Более точно итоговая величина суммарного диффузионного потенциала, который создаётся утечкой нескольких видов ионов, рассчитывается по формуле Гольдмана-Ходжкина-Катца. В ней учтено, что потенциал покоя зависит от трех факторов: (1) полярности электрического заряда каждого иона; (2) проницаемости мембраны Р для каждого иона; (3) [концентраций соответствующих ионов] внутри (внутр) и снаружи мембраны (внеш). Для мембраны аксона кальмара в покое отношение проводимостей РK : PNa :PCl = 1 : 0,04 : 0,45 [5].
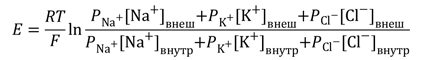
Заключение
Итак, поте нциал покоя состоит из двух частей:
- −10 мВ, которые получаются от «несимметричной» работы мембранного насоса-обменника (ведь он больше выкачивает из клетки положительных зарядов (Na + ), чем закачивает обратно с калием).
- Вторая часть — это всё время утекающий из клетки калий, уносящий положительные заряды. Его вклад — основной: −60 мВ. В сумме это и дает искомые −70 мВ.
Что интересно, калий перестанет выходить из клетки (точнее, его вход и выход уравниваются) только при уровне отрицательности клетки −90 мВ. В этом случае сравняются химические и электрические силы, проталкивающие калий через мембрану, но направляющие его в противоположные стороны. Но этому мешает постоянно подтекающий в клетку натрий, который несёт с собой положительные заряды и уменьшает отрицательность, за которую «борется» калий. И в итоге в клетке поддерживается равновесное состояние на уровне −70 мВ.
Вот теперь мембранный потенциал покоя окончательно сформирован.
Схема работы Na + /K + -АТФазы наглядно иллюстрирует «несимметричный» обмен Na + на K + : выкачивание избыточного «плюса» в каждом цикле работы фермента приводит к отрицательному заряжению внутренней поверхности мембраны. Чего в этом ролике не сказано, так это того, что АТФаза ответственна за менее чем 20% потенциала покоя (−10 мВ): оставшаяся «отрицательность» (−60 мВ) появляется за счет выхода из клетки через «калиевые каналы утечки» ионов K + , стремящихся выровнять свою концентрацию внутри клетки и вне нее.
Как найти потенциал капли
Мембранный потенциал и электрохимический градиент
• Мембранный потенциал возникает за счет электрохимического градиента, который существует по обеим сторонам мембраны, селективно проницаемой для ионов
• Величина мембранного потенциала как функции концентрации ионов рассчитывается по уравнению Нернста
• В клетке поддерживается отрицательное значение мембранного потенциала покоя. При этом внутренняя среда клетки, по сравнению с внешней, характеризуется несколько большим отрицательным зарядом
• Существование мембранного потенциала является необходимым условием генерации электрических сигналов, а также направленного транспорта ионов через мембрану
Важным свойством клеток является способность поддерживать такие внутриклеточные концентрации метаболитов, которые существенно отличаются от их содержания во внеклеточной среде. В случае ионов, различия в их концентрации по обеим сторонам мембраны приводят к различиям в электрическом заряде: внутриклеточная среда заряжена несколько более отрицательно, чем среда снаружи клетки. Совместное действие разности зарядов и концентраций проводит к возникновению электрохимического градиента. Электрохимический градиент поддерживается за счет действия селективных каналов и белков переносчиков в плазматической мембране.
Для того чтобы понять, каким образом возникает электрохимический градиент, вначале рассмотрим простой случай, когда мембрана оказывается проницаемой только для одного вида ионов. На рисунке ниже представлены два компартмента, А и В, разделенные тонкой мембраной. Эти компартменты содержат раствор КС1 разной концентрации. В растворе хлорид калия диссоциирован на гидратированные ионы К+ и Cl-. Поскольку оба компартмента содержат эквимолярные концентрации ионов, то каждый обладает нейтральным зарядом.
Если бы мембрана была непроницаема для ионов, то величина ее электрического потенциала, измеренная с помощью вольтметра, равнялась бы нулю.
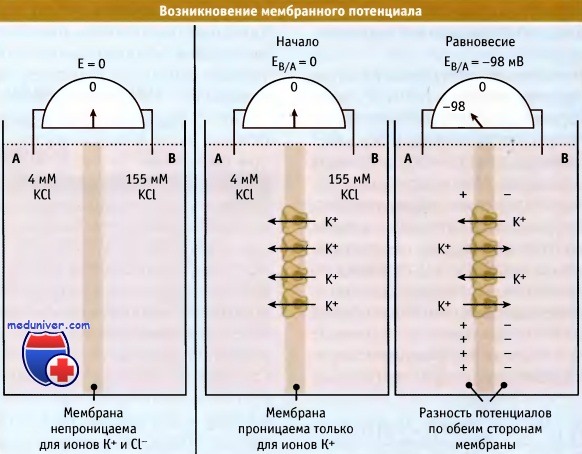 Селективное передвижение ионов через мембрану вызывает изменение мембранного потенциала.
Селективное передвижение ионов через мембрану вызывает изменение мембранного потенциала.
Теперь рассмотрим случай, когда мембрана проницаема только для ионов калия (например, когда в мембране находятся К+-каналы). Диффузия растворенных веществ по градиенту концентрации является энергетически выгодным процессом (выражается в виде отрицательной величины разности энергии AG). Поэтому ионы К+ будут диффундировать в сторону более низкой их концентрации, т. е. из компартмента В в компартмент А. При этом распределение заряда на мембране будет меняться. По мере накопления в компартменте А положительно заряженных ионов, возрастают силы отталкивания между ними. Эти силы затрудняют переход ионов К+ в компартмент А.
Когда в системе достигается электрохимическое равновесие, градиенты концентрации и электрических зарядов взаимно уравновешиваются, и движение ионов К+ через мембрану прекращается. При этом транспорт ионов К+ из одного компартмента сдерживается их транспортом из другого компартмента.
Однако в компартменте А содержится больше положительно заряженных ионов, чем в компартменте В. Этот избыток ионов К+ (в компартменте А) взаимодействует с избытком ионов Cl- (в компартменте В) через тонкую мембрану, в результате чего по обеим ее сторонам выстраиваются электрические заряды. Разница зарядов по обеим сторонам мембраны выражается в виде разности потенциалов и называется мембранный потенциал. Равновесный (мембранный) потенциал компартмента В по отношению к компартменту А имеет отрицательное значение.
Этот пример иллюстрирует необходимость наличия двух условий, необходимых для возникновения мембранного потенциала клетки, не равного нулю:
• различные концентрации ионов по обеим сторонам мембраны, которые приводят к разделению зарядов и
• мембрана, обладающая селективной проницаемостью по крайней мере к одному виду ионов.
Поэтому величина мембранного потенциала является функцией концентрации ионов. В состоянии равновесия эту функцию для ионов X можно выразить количественно с помощью уравнения Нернста:
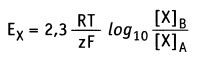
• Е — равновесный потенциал (в вольтах)
• R — универсальная газовая постоянная (2 кал моль -1 К -1 )
• Т — абсолютная температура (К; 37 °С = 307,5 К)
• z — валентность ионов (электрический заряд)
• F — число Фарадея (2,3 х 10 4 кал вольт -1 моль -1 )
• [Х]А — концентрация свободных ионов X в компартменте А
• [Х]в — концентрация свободных ионов X в компартменте В
В формировании мембранного потенциала в клетках животных, главным образом, участвуют ионы К+, Na+ и Cl-. Ионы Са2+ и Mg2+ в меньшей степени участвуют в формировании мембранного потенциала покоя. Плазматическая мембрана обладает селективной проницаемостью к перечисленным ионам (т. е. мембрана содержит ионные каналы, селективные к каждому типу ионов). Это обстоятельство, а также мембранная проницаемость (Р) для каждого иона учитывается в уравнении Гольдмана-Ходжкина-Каца, которое представляет собой расширенную форму уравнения Нернста.
Для основных ионов это уравнение выражает мембранный потенциал как функцию их проницаемости и концентрации внутри (i) и снаружи (о) клетки:
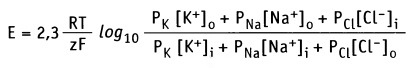
Величина отрицательного мембранного потенциала покоя зависит от типа клеток и колеблется от -200 мВ до -20 мВ. В клетках млекопитающих мембранный потенциал покоя в основном создается при работе К+-каналов и ионного насоса, который называется Na+/К+-АТФаза. Основной вклад в формирование отрицательного мембранного потенциала вносит небольшой поток ионов К+ через плазматическую мембрану. Этот поток осуществляется через К+-каналы, лишенные воротного механизма (т. н. калиевые каналы покоя).
В отличие от большинства других К+-каналов, которым необходим сигнал для открытия, эти каналы в клетке, обладающей определенным потенциалом покоя, открыты постоянно. В покоящейся клетке также открыты несколько каналов для других ионов. Движение ионов К+ из клетки, по направлению электрохимического градиента, помогает клеточному содержимому поддерживать отрицательный заряд. Пока мы не знаем всех источников ионов калия, которые участвуют в этом процессе. В некоторых клетках, например у растений и бактерий, а также в митохондриях, мембранный потенциал покоя создается за счет градиента протонов, а не ионов К+.
Для того чтобы происходила диффузия ионов К+ из клетки через К+-каналы, их концентрация в клетке должна быть выше, чем в окружающей среде. Градиент концентрации создается в результате работы Na+/К+-АТФа-зы, которая закачивает в клетку два иона калия на каждые три иона натрия, которые этот ионный насос удаляет из клетки. Поэтому насос функционирует как генератор заряда: удаляется больше электрических зарядов, чем привносится к клетку. Таким образом, наряду с K+-каналами, лишенными воротного механизма, Na+/К+-АТФазы участвуют в создании отрицательного внутриклеточного потенциала. Если происходит инактивация Na+/K+-АТФаз, то концентрации ионов Na+ и К+ по обе стороны мембраны уравниваются. Это происходит потому, что липидный бислой очень плохо пропускает ионы. Иными словами, без прохождения первичных процессов активного транспорта с участием Na+/К+-АТФаз значение мембранного потенциала равнялось бы нулю.
Мембранный потенциал покоящейся клетки представляет собой довольно постоянную величину. Однако при связывании лигандов, механическом стрессе или при изменении электрического заряда происходит открытие специфических ионных каналов, и мембранный потенциал изменяется. Если ионные каналы находятся под контролем электрического заряда, то изменения мембранного потенциала влияют на прохождение через них ионов. Открытие и закрытие канала контролируются воротным механизмом (гейтингом). Мембранный потенциал зависит от тех ионов, для которых каналы в основном, открыты. Например, при открытии Na+- или Са2+-каналов происходит деполяризация мембраны.
При этом соответствующие ионы начинают поступать в клетку в направлении их электрохимического градиента. Это приводит к тому, что мембранный потенциал становится более положительным. Напротив, при реполяризации мембраны (гиперполяризации) потенциал становится еще более отрицательным. Это происходит при открытии калиевых каналов и выходе из клетки ионов К+ в направлении градиента, что и влечет за собой увеличение отрицательного мембранного потенциала. Движение ионов по ионным каналам происходит быстро и исчисляется миллисекундами. Для изменения мембранного потенциала достаточны лишь незначительные различия в концентрации ионов по сторонам мембраны, и основная концентрация их в клетке не меняется.
Поток лишь 10 -12 моль К+ через 1 см2 мембраны приводит к ее быстрой гиперполяризации и к установлению мембранного потенциала, равного -100 мВ. Локальное передвижение относительно небольших зарядов через мембрану позволяет цитозолю и внеклеточной среде оставаться электрически нейтральными и сводит к минимуму электрическое отталкивание зарядов.
С энергетической точки зрения, мембранный потенциал представляет собой некий энергетический резервуар, энергию которого можно использовать для выполнения определенной работы. По расположению отрицательно заряженных ионов в цитозоле и положительно заряженных на наружной стороне мембраны, клетка напоминает электрический конденсатор или батарею, т. е. приспособление, способное сохранять электрическую энергию и служить ее источником. Энергия высвобождается в виде ионов, мигрирующих по направлению их электрохимического градиента, и может использоваться в процессах транспорта других ионов или метаболитов против градиента концентрации.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
2017-08-20 ![]()
Капля ртути, заряженная до потенциала $phi_{0}$, распадается на $N$ одинаковых капель с одинаковыми зарядами. Капли разлетаются на большое расстояние друг от друга. Определите потенциал $phi$ каждой из образовавшихся капель.
Решение:
Потенциал заряженной капли (проводящего шара) $phi_{0} = k frac{Q}{R}$, заряд каждой, из маленьких капель $q = frac{Q}{N}$. Радиус $r$ маленьких капель найдем из условия неизменности объема ртути:
$frac{4 pi R^{3}}{3} = N frac{4 pi r^{3}}{3}$,
откуда $r = frac{R}{N^{1/3}}$. Тогда потенциал образовавшихся капель составит $phi = k frac{q}{r} =k frac{Q}{N^{2/3}R} = frac{ phi_{0}}{N^{2/5}}$, т. е. уменьшается. Отметим, что этот ответ справедлив только в том случае, когда образовавшиеся капли разлетятся на большое расстояние (чтобы можно было пренебречь энергией их взаимодействия).
Ответ: $phi = frac{ phi_{0}}{ N^{2/3}}$.
Решение. Потенциалы φ «маленькой» и φ1 «большой» капли равны соответственно
[ varphi =kcdot frac{q}{r};,,,,,{{varphi }_{1}}=kcdot frac{4cdot q}{R},,(1) ]
Объемы капель
[ V=frac{4}{3}cdot pi cdot {{r}^{3}};,,,,,{{V}_{1}}=frac{4}{3}cdot pi cdot {{R}^{3}},,,,(2) ]
С учетом того, что V1 = 4·V из (2)
[ R=rcdot sqrt[3]{4} ]
Решим уравнения (1). Например
[ begin{align}
& frac{{{varphi }_{1}}}{varphi }=frac{kcdot 4cdot q}{R}cdot frac{r}{kcdot q}=frac{4cdot r}{R}=frac{4}{sqrt[3]{4}}; \
& ,,,,,,{{varphi }_{1}}=varphi cdot frac{4}{sqrt[3]{4}}=varphi cdot sqrt[3]{{{4}^{2}}} \
end{align} ]
φ1 = 25.2 В
В
насыщенном паре образовалась капля
жидкости с коэффициентом поверхностного
натяжения
радиусом R.
Получим условия увеличения и уменьшения
размера капли. Аналогичная задача
возникает для квантовой точки, образующейся
в насыщенном растворе.
При
образовании капли происходит переход
N
частиц пара в жидкость. Для этого
химический потенциал пара
![]() должен превышать химический потенциал
должен превышать химический потенциал
жидкости![]()
![]() .
.
При изотермическом и изохорическом
образовании капли система равновесная
при минимуме свободной энергии![]() .
.
Из (2.67) с учетом энергии поверхностного
натяжения жидкости![]() получаем
получаем
![]() ,
,
где
S
– площадь поверхности жидкости.
При
образовании капли увеличивается площадь
поверхности жидкости на
![]() ,
,
гдеR
– радиус капли. Изменение свободной
энергии
![]() ,
,
(П.7.15)
где
 –число частиц
–число частиц
жидкости в капле; v
– объем частицы.
В
равновесной системе размер капли не
меняется. Равновесие соответствует
экстремуму свободной энергии
 .
.
Используем
![]() ,
,
получаем
равновесный
радиус капли
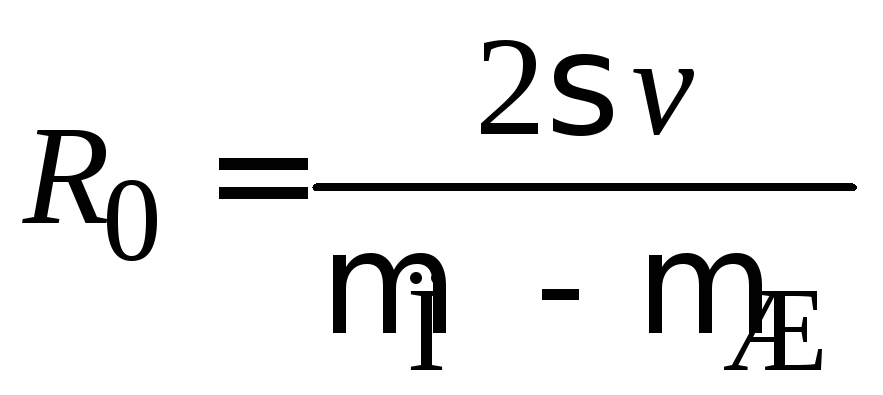 .
.
(П.7.16)
С
учетом

выполняется
 .
.
Следовательно,
свободная энергия максимальна. Устойчивое
равновесие соответствует минимуму
свободной энергии, поэтому состояние
капли неустойчиво:
при
![]() капля увеличивается; при
капля увеличивается; при![]() капля испаряется.
капля испаряется.
Как изменится состояние капли, если она
образуется вокруг иона?
7. Заряженная капля в насыщенном паре
Ион
радиусом
![]() имеет зарядq,
имеет зарядq,
его окружает пар c
химическим потенциалом
![]() .
.
Вокруг иона образовалась капля жидкости
радиусомR
c
химическим потенциалом
![]() ,
,
причем![]() .
.
Найдем изменение свободной энергии при
образовании капли.
Используем
обозначения и результаты примера 4.
Дополнительно учитываем энергию
электрического поля капли
 ,
,
где
 –плотность
–плотность
электрической энергии;
![]() –объем шарового
–объем шарового
слоя радиусом r
толщиной dr;
– диэлектрическая
проницаемость жидкости;
 –напряженность
–напряженность
поля внутри капли при
![]() ;
;
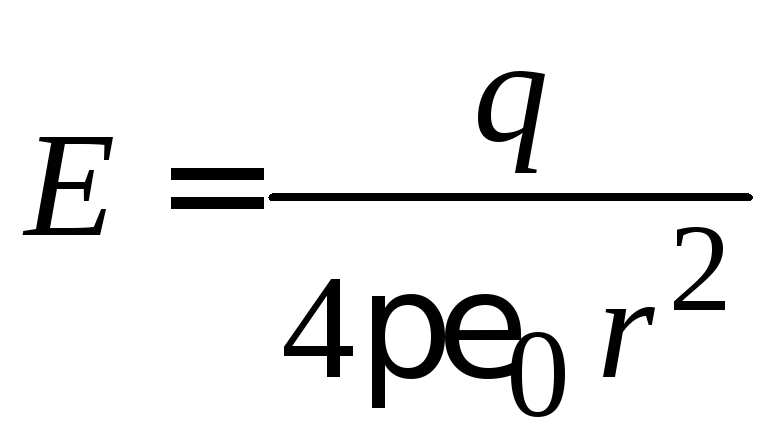 –напряженность
–напряженность
поля вне капли.
К
вкладу без поля (П.7.15)
![]()
Добавляем
энергию поля внутри и вне капли

= .
.
При
достаточно большом заряде q
и малом размере иона
![]() главную роль играет третье слагаемое.
главную роль играет третье слагаемое.
Оно отрицательное и уменьшается с
увеличением радиуса каплиR.
При уменьшении свободной энергии
возрастает устойчивость системы. В
результате заряженная
капля увеличивается даже в ненасыщенном
паре.
Если
капля получает заряд за счет эмиссии
электронов, то, как показал Джон У. Рэлей
в 1882 г., при достаточно малом размере R0
~ 50 мкм, называемом пределом
Рэлея,
капля становится неустойчивой из-за
преобладания сил электростатического
отталкивания над поверхностным
натяжением. Капля вытягивается, из ее
концов выбрасывается заряд, переносимый
струями жидкости.
Условия применимости классической статистической физики
На
рубеже XIX–XX
вв. делались попытки применить классическую
статистическую физику для описания
электромагнитного теплового излучения
в полости, электронного газа в металле,
тепловых свойств кристалла. Оказалось,
что в этих областях следствия теории
противоречат опыту. Рассмотрим примеры.
Теплоемкость
твердых тел.
Металл состоит из кристаллической
решетки ионов, потерявших свои валентные
электроны, и электронов, ставших
свободными. Для моля одновалентного
металла количество ионов равно количеству
свободных электронов и равно числу
Авогадро
![]() .
.
Теплоемкость металла
![]()
содержит
вклады электронного газа
![]() и ионов решетки
и ионов решетки![]() .
.
Свободный электрон имеет 3 степени
свободы и по теореме о распределении
энергии по степеням свободы его средняя
энергия![]() .
.
Внутренняя энергия моля и молярная
теплоемкость электронного газа
![]() ,
,
![]() ,
,
где
универсальная газовая постоянная
![]() .
.
Ион решетки имеет 3 колебательные степени
свободы, на каждую приходится тепловая
энергия![]() ,
,
тогда средняя энергия иона, внутренняя
энергия моля и молярная теплоемкость
кристаллической решетки
![]() ,
,
![]() ,
,![]() .
.
Для
молярной теплоемкости металла получаем
![]() ,
,
для
диэлектрика, не имеющего свободных
электронов:
![]() .
.
Эксперименты
при нормальной и более высокой температурах
подтверждают закон Дюлонга и Пти (П.4.7)
![]()
как
для металлов, так и для диэлектриков.
Следовательно, электронный
газ не дает вклада в теплоемкость
металла.
При температуре, существенно меньшей
нормальной, эксперимент обнаруживает
зависимость теплоемкости от температуры
– для диэлектриков
![]() ;
;
для металлов![]() .
.
Для объяснения этих результатов
необходимо учитывать квантовые свойства
микрочастиц.
Магнетизм
системы зарядов.
Согласно теореме Бора
– ван Лёвен, рассмотренной в Примере
3.10, система
зарядов не проявляет магнитных свойств,
что противоречит свойствам магнетизма
ряда металлов.
Ограничения
на параметры системы.
Требуется очертить рамки, в пределах
которых применима классическая физика
и несущественны квантовые
свойства, главными из которых являются:
1.
Интерференция и дифракция частиц,
приводящие к квантованию энергии системы
и к дискретности фазового пространства.
Волновые свойства частицы описываются
длиной волны де Бройля
![]() ,
,
и условием квантования Бора–Зоммерфельда на траектории
на траектории
![]() ;
;
2.
Вырождение состояний по энергии;
3.
Тождественность микрочастиц и принцип
запрета Паули для фермионов.
Ограничим
области параметров системы, для которых
применима классическая статистическая
физика.
Высокие
температуры.
Для частицы в кубическом ящике со
стороной L
условие квантования Бора–Зоммерфельда
допускает дискретные значения импульса
![]() .
.
Энергия частицы квантуется .
.
Дискретность спектра несущественна,
если расстояние между уровнями гораздо
меньше тепловой энергии
 .
.
(2.190)
Для
![]() и массы электрона получаем
и массы электрона получаем![]() ,
,
что равно тепловой энергии![]() при температуре
при температуре![]() .Классическая
.Классическая
теория выполняется для частиц, находящихся
в макроскопическом объеме при относительно
высокой температуре, если масса частицы
не слишком мала.
Малая
концентрация частиц.
Длина волны
де Бройля несущественна при
большом
расстоянии r
между частицами
![]() .
.
Для
наиболее вероятной скорости частицы
газа (2.124)
![]() ,
,
импульс![]() и длина волны де Бройля
и длина волны де Бройля
 .
.
Среднее
расстояние
![]() междуN
междуN
частицами газа выражаем через объем
сосуда V
и концентрация
![]() ,
,
находим![]() ,
,
![]() .
.
Получаем
условие
![]() .
.
(2.191)
Классическая
теория идеального газа выполняется при
достаточно малой концентрации, высокой
температуре и не слишком малой массе
частицы.
Как следует из (2.162), в
области применимости классической
теории химический
потенциал газа отрицательный.
Для
гелия при нормальных условиях
![]() ,
,
![]() ,
,
![]() ,
,
![]() ,
,![]() ,
,
выполняется
![]() и классическая физика применима.
и классическая физика применима.
Для
электронов металла при нормальной
температуре
![]() ,
,
![]() ,
,![]() .
.
Классическая
физика не применима для электронного
газа в металле.
Для
полупроводников длина волны де Бройля
электрона в нанометрах дана в таблице.
|
Ge |
Si |
GaAs |
InSb |
|
|
|
12 |
15,4 |
58 |
134 |
|
|
6 |
7,7 |
29 |
67 |
В
узкозонных полупроводниках с шириной
запрещенной зоны
![]() ,
,
напримерInSb
и InAs,
концентрация электронов проводимости
велика,
![]() ,
,
условие классического описания![]() нарушается.
нарушается.
Большой
объем фазового пространства,
занимаемый узлом кристаллической
решетки, по сравнению с постоянной
Планка
![]() .
.
(2.192)
Флуктуации
импульса и координаты выражаем через
параметры системы. По теореме о
распределении энергии по степеням
свободы средняя кинетическая энергия
одномерного движения узла кристаллической
решетки
![]() ,
,
тогда
![]() .
.
С учетом![]() тепловая флуктуация проекции импульса
тепловая флуктуация проекции импульса
![]() .
.
Для
потенциальной энергии упругих колебаний
с частотой ω аналогично находим
![]() ,
,
 .
.
Чем
прочнее кристалл, тем больше частота –
для меди
![]() ,
,
для алмаза![]() .
.
Тепловая флуктуация координаты узла с
учетом![]() равна
равна
 .
.
В результате
![]() .
.
Условие
(2.192) получает вид
![]() .
.
(2.193)
Чем
прочнее кристалл и ниже температура,
тем меньше согласие с классической
теорией.
Классическая физика не применима для
прочных кристаллов при низкой температуре.
Для
систем, нарушающих хотя бы одно из
полученных условий, необходимо
использовать квантовую статистическую
физику.
Соседние файлы в папке сгф
- #
- #
- #
поделиться знаниями или
запомнить страничку
- Все категории
-
экономические
43,662 -
гуманитарные
33,654 -
юридические
17,917 -
школьный раздел
611,978 -
разное
16,905
Популярное на сайте:
Как быстро выучить стихотворение наизусть? Запоминание стихов является стандартным заданием во многих школах.
Как научится читать по диагонали? Скорость чтения зависит от скорости восприятия каждого отдельного слова в тексте.
Как быстро и эффективно исправить почерк? Люди часто предполагают, что каллиграфия и почерк являются синонимами, но это не так.
Как научится говорить грамотно и правильно? Общение на хорошем, уверенном и естественном русском языке является достижимой целью.