Динамика
популяций — это процесс изменения ее
основных биологических характеристик
во времени.
Динамика
популяций является одним из наиболее
значимых биологических и экологических
явлений. Можно сказать, что жизнь
популяции проявляется в ее динамике,
так как популяция не может существовать
без постоянных изменений, за счет которых
она как бы приспосабливается к постоянно
происходящим изменениям внешних условий.
Основными
динамическими показателями популяций
являются рождаемость, смертность и
скорость роста популяций.
2.3.1. Рождаемость, смертность, миграции особей.
Динамика
численности и плотности популяций
находится в тесной зависимости от
рождаемости и смертности, а также от
способности слагающих их особей совершать
миграционные процессы.
Рождаемость
— это показатель, характеризующий
скорость увеличения численности
популяции в результате размножения, т.
е. показатель, характеризующий скорость
размножения.
Различают
абсолютную
и удельную рождаемость
Абсолютная
или общая рождаемость (Р) определяется
как число особей (∆Nп)
родившихся в популяции за определенный
промежуток времени (∆t):
Р
= ∆Nп
/ ∆t
Удельная
рождаемость (b)
– это отношение особей (∆Nп),
родившихся за определенный промежуток
времени (∆t),
к числу всех особей популяции (N):
b
= ∆Nп
/ ∆t
·
N
Этот показатель
применяют для сравнения между собой
рождаемости в популяциях, имеющих разную
численность.
Например:
если в популяции
имеется 500 особей способных к размножению
(N
=500) и в течение 10 дней (∆t
) родилось
50 новых особей (∆Nп
=50),
то абсолютная или
общая рождаемость составит: Р
= ∆Nп
/ ∆t
= 50/10 = 5,
а удельная
рождаемость: b
= ∆Nп
/ ∆t
·
N
= 50/ 10 · 500
= 0,01 или 1%
Различают
максимальную
и фактическую рождаемость.
Максимальная
(физиологическая) рождаемость — это
теоретический максимум скорости рождения
новых особей в идеальных условиях при
отсутствии лимитирующих факторов среды.
Максимальная
рождаемость постоянна для данной
популяции, и определяется только
физиологическими особенностями
организмов (физиологической плодовитостью
самок).
В живых
организмах заложена огромная потенциальная
возможность к размножению, поэтому при
неограниченных ресурсах и идеальных
природных условиях виды реализуют
максимальную рождаемость.
Для каждой
популяции характерен биотический
(или репродуктивный) потенциал, т.е.
скорость увеличения численности особей
популяции при отсутствии лимитирующих
факторов среды.
Биотический
(или репродуктивный) потенциал
устанавливается теоретически как
скорость, с которой при беспрерывном
размножении (возможном только теоретически
при идеальных экологических условиях
существования) особи определенного
вида могут покрыть земной шар равномерным
слоем.
Например:
— для слонов эта
скорость составляет 0,3 м/с;
— для некоторых
микроорганизмов — сотни метров в секунду.
Это означает, что
при отсутствии лимитирующих факторов
среды, такие быстроразмножающиеся виды,
как бактерии, насекомые, мыши и некоторые
рыбы, могут увеличить свою численность
до заселения всей поверхности Земли за
короткий промежуток времени.
Подсчитано,
что:
— бактерии делятся
каждые 20 минут. При таком темпе одна
клетка за 36 часов дает потомство,
которое может покрыть сплошным слоем
всю нашу планету;
— один одуванчик
менее чем за 10 лет способен заселить
своими потомками земной шар, если все
семена прорастут.
В действительности
же такая громадная рождаемость организмов
никогда не реализуется и фактическая
рождаемость значительно ниже максимальной.
Фактическая
(или экологическая) рождаемость
характеризует скорость размножения
популяции при фактически сложившихся
условиях жизни, т.е. в конкретных
экологических условиях среды.
Фактическая
рождаемость зависит от многих абиотических
и
биотических
факторов среды
(неблагоприятные климатические условия,
потеря или ухудшение естественной среды
обитания из-за стихийных бедствий —
засуха, землетрясения, ураганы, пожары,
наводнения или в результате человеческой
деятельности, недостаток пиши,
внутривидовая и межвидовая конкуренция,
хищничество, паразитизм, болезнетворные
агенты и т.д.), а
также от возраста и состояния популяции.
Смертность –
величина, обратная рождаемости.
Смертность
— это показатель, характеризующий
скорость процесса снижения численности
популяции в результате гибели особей.
Различают
общую и
удельную смертность.
Общая
смертность (С) определяется как число
погибших в популяции особей (∆Nm)
за определенный промежуток времени
(∆t):
С
= ∆Nm
/ ∆t
Удельная
смертность (d)
– это отношение особей (∆Nm),
погибших за определенный промежуток
времени (∆t),
к числу всех особей популяции (N):
d
= ∆Nm
/ ∆t
·
N
Различают
минимальную
и фактическую смертность.
Минимальная
(физиологическая) смертность — это
минимально возможная величина смертности,
проявляющаяся даже в идеальных условиях
среды как результат физиологической
старости особей.
В этом случае особи
реализуют весь генетически определенный
цикл жизнедеятельности организма.
В конкретных
экологических условиях фактическая
(экологическая)
смертность,
как правило, выше
минимального уровня, так как под влиянием
абиотических и биотических факторов
среды создаются дополнительные причины
гибели организмов.
Смертность
влияет на продолжительность жизни
организмов в популяции и тем самым на
ее возрастной состав.
В зависимости
от возрастного состава популяций
различают три
типа смертности.
Первый тип
смертности характеризуется повышенной
гибелью взрослых, главным образом,
старых особей.
Этот тип
смертности наблюдается:
— у насекомых,
личинки которых обитают в почве, воде,
древесине или в других местах с
благоприятными условиями;
— у большинства
крупных млекопитающих, которые заботятся
о своем потомстве длительное время;
— у людей с
правильным питанием и здоровым образом
жизни.
Ко второму
типу относится смертность, одинаковая
для всех возрастных групп.
Такая смертность
встречается очень редко и только у
популяций, постоянно находящихся в
экстремальных условиях:
— наблюдается у
некоторых певчих птиц, рептилий и мелких
млекопитающих.
Третий тип
смертности отличается повышенной
гибелью особей на ранних стадиях своего
развития.
Этот тип
смертности свойственен большинству
видов морских животных и растений:
— морские виды
животных (устрицы или другие двустворчатые
моллюски, а также рыбы) дают огромное
потомство, но только некоторые особи
доживают до половозрелого возраста;
— растения,
размножающиеся благодаря рассеиванию
большого числа семян, и у которых
максимальная гибель происходит в стадии
прорастания семян и всходов.
Обычно смертность
представляется графически в виде “кривых
выживания”,
которые
выражают зависимость числа выживших
из 100 или 1000 особей от их возраста.
Соответственно
трем типам смертности было получено
три типа
кривых выживания (рис.1):

Рис.1.
Три типа кривых выживания
1 кривая
выживания
характеризуется слабым понижением
смертности до возраста естественной
(“физиологической”) смертности с
последующим резким падением, отражающим
вымирание особей, достигших этого
критического возраста;
2 кривая выживания
— это идеальная диагональная кривая
выживания — представляется в виде
диагонально снижающей прямой линии и
характеризуется равномерным распределением
смертности по возрастам;
3 кривая выживания
демонстрирует резкое падение смертности
в области младших возрастов, которое
вскоре сменяется постепенным относительно
равномерным понижением смертности
организмов, переживших “критический”
возраст.
Существуют также
внутривидовые различия в кривых
выживания. Причины могут быть разные и
нередко связаны с полом. Так, женщины
живут дольше мужчин, поэтому кривая
выживания для мужчин по сравнению с
аналогичной кривой для женщин менее
выпуклая.
Кроме того,
2 кривая выживания, построенная по
надписям на надгробиях людей, живших в
Древнем Риме в 1-1У веках н.э., характерна
только для древних римлян, а для других
народностей – 1 кривая выживания.
Анализ кривых
выживания и смертности в разных возрастных
группах открывает возможность расчета
ожидаемой продолжительности жизни
особей данной возрастной группы, что
имеет большое значение в прикладных
экологических исследованиях.
На динамику
численности и плотности популяций
большое влияние оказывают миграции
особей, которые
происходят при перемещении особей из
одного местообитания (эмиграция)
в другое
(иммиграция).
Миграции
— закономерные перемещения живых
организмов между разными средами
обитания, пространственно отстоящими
друг от друга. Происходят по разным
причинам:
— изменение условий
существования в местах обитания;
— изменений
требований живых организмов к этим
условиям на разных стадиях развития.
В основном
миграции совершаются при подрастании
молодняка и его расселении.
Наиболее
интенсивно расселение происходит при
несбалансированных процессах рождаемости
и смертности в популяции. В результате
миграции происходит обмен особями
популяций, в результате чего устраняется
избыток особей в одной популяции и
компенсируется в другой.
Миграции
(суточные, сезонные) позволяют организмам
использовать оптимальные условия среды
в таких местах, где их постоянное
проживание невозможно. Они приводят к
освоению новых биотопов, расширению
общего ареала вида, к обмену особей
между популяциями, увеличивают единство
и общую устойчивость вида, способствуют
успеху в борьбе за существование.
В разных
популяциях рождаемость, смертность и
миграции особей различаются. Они являются
специфичными для каждого вида и зависят
от множества причин, обусловливающих
особенности рождаемости, смертности и
подвижности особей.
Таким образом,
рассмотренные четыре переменных фактора
— рождаемость,
смертность, иммиграция и эмиграция
определяют
изменение численности популяции.
Особи появляются
в популяции благодаря рождению и
иммиграции, а исчезают в результате
смерти и эмиграции:
изменение
численности популяции = (рождаемость +
иммиграция) — (смертность + эмиграция).
При
отсутствии миграции изменение численности
популяции зависит от соотношения
величины рождаемости и смертности.
Соседние файлы в папке Тема 3. Вид и популяция
- #
- #
- #
- #
- #
- #
17.03.2016146.43 Кб38~WRL0711.tmp
- #
А.С. Степановских
Экология. Учебник для вузов
М.: ЮНИТИ-ДАНА, 2001. — 703 с.
Содержание статьи:
- 1 9. Структура и динамика популяций
- 1.1 9.4. Рождаемость и смертность
9. Структура и динамика популяций
9.4. Рождаемость и смертность
Динамика численности и плотности популяций находится в тесной зависимости от рождаемости или плодовитости и смертности.
Рождаемость — это способность популяции к увеличению численности. Характеризует частоту появления новых особей в популяции. Различают рождаемость абсолютную и удельную. Абсолютная (общая) рождаемость — число новых особей (DNn), появившихся за единицу времени (Dt). Удельная рождаемость выражается в числе особей на особь в единицу времени:
![]() . (9.2)
. (9.2)
Так, для популяций человека как показатель удельной рождаемости используют число детей, родившихся в год на 1000 человек. В живых организмах заложена огромная возможность к размножению и подтверждается правилом максимальной рождаемости (воспроизводства): в популяции имеется тенденция к образованию теоретически максимально возможного количества новых особей. Оно достигается в идеальных условиях, когда отсутствуют лимитирующие экологические факторы и размножение ограничено лишь физиологическими особенностями вида. Например, один одуванчик менее чем за 10 лет способен заселить своими потомками земной шар, если все семена прорастут. Другой пример. Бактерии делятся каждые 20 мин. При таком темпе одна клетка за 36 ч может дать потомство, которое покроет сплошным слоем всю нашу планету. Обычно же существует экологическая или реализуемая рождаемость, возникающая в обычных или специфических условиях среды. Средняя величина плодовитости выработана исторически как приспособление, которое обеспечивает пополнение убыли популяций. Естественно, что у менее приспособленных видов к неблагоприятным условиям высокая смертность в молодом (личиночном) возрасте компенсируется значительной плодовитостью.
Среди насекомых самая высокая плодовитость у растительноядных форм, а низкая — у хищников и паразитов. В благоприятных условиях плодовитость, как правило, низкая. Характер плодовитости зависит и от скорости полового созревания, числа генераций в течение сезона, от состояния в популяции самок и самцов. Если вид размножается с большой скоростью и чутко реагирует на изменения условий среды, то численность популяций его быстро и существенно изменяется. Это относится к многим насекомым и мышевидным грызунам. Таким образом, максимальная рождаемость или плодовитость является константой, определяемой расчетным путем, например, умножением среднего числа гнезд, которое способна построить самка птицы за год, на такое же число яиц, которые она может отложить в наиболее благоприятную часть сезона года. Максимальная рождаемость — тот предел, который характерен для скоростей увеличения числа особей в популяции. Правило максимальной рождаемости (воспроизводства) есть частный случай закона максимума биогенной энергии (энтропии) В. И. Вернадского — Э. С. Бауэра.
Численность и плотность популяции зависит и от ее смертности. Смертность популяции — это количество особей, погибших за определенный период. Абсолютная (общая) смертность — это число особей, погибших в единицу времени (DNm).
Удельная смертность (d) выражается отношением абсолютной смертности к численности популяции:
![]()
Абсолютная и удельная смертность характеризуют скорость убывания численности популяции вследствие гибели особей от хищников, болезней, старости и т. д.
Различают три типа смертности. Первый тип смертности характеризуется одинаковой смертностью во всех возрастах. Выражается экспоненциальной кривой (убывающей геометрической прогрессии). Данный тип смертности встречается редко и только у популяций, которые постоянно находятся в оптимальных условиях.
Второй тип смертности характеризуется повышенной гибелью особей на ранних стадиях развития и свойствен большинству растений и животных. Максимальная гибель животных происходит в личиночной фазе или в молодом возрасте, у многих растений — в стадии произрастания семян и всходов. У насекомых до взрослых особей доживает 0,3—0,5% отложенных яиц, у многих рыб — 1—2% количества выметанной икры.
Третий тип смертности отличается повышенной гибелью взрослых, в первую очередь старых, особей. Отличается он у насекомых, личинки которых обитают в почве, воде, древесине, а также в других местах с благоприятными условиями. В экологии широкое распространение получило графическое построение «кривых выживания» (рис. 9.3).
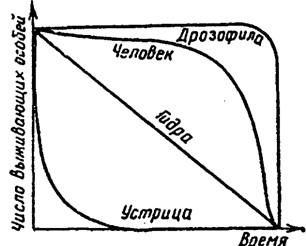
Рис. 9.3. Различные типы кривых выживания
Откладывая по оси абсцисс продолжительность жизни в процентах от общей продолжительности жизни, можно сравнивать кривые выживания организмов, продолжительность жизни которых имеет значительные различия. На основании таких кривых можно определить периоды, в течение которых тот или иной вид особенно уязвим. Поскольку смертность подвержена более резким колебаниям и больше зависит от факторов окружающей среды, чем рождаемость, она играет главную роль в регулировании численности популяции.
Смертность — способность популяции снижать численность особей в популяции за счет их гибели, причем причина гибели может быть любой (поедание хищниками, гибель от болезней, недостаток корма или естественное старение). Смертность по характеру влияния на численность природных популяций является свойством, противоположным рождаемости. Смертность в идеальных условиях, когда нет лимитирующих факторов, является минимальной и зависит от потенциальной генетически детерминированной продолжительности жизни особей. В реальных условиях среды наблюдается реализованная, или экологическая смертность. Количественно она выражается числом особей, умерших за единицу времени — абсолютная смертность. Показатель абсолютной смертности, как и в случае с абсолютной рождаемостью, нельзя использовать для сравнения популяций с разной начальной численностью по смертности. Для этого абсолютную смертность нужно соотнести с начальной численностью и определить удельную смертность, или количество умерших особей за единицу времени в расчете на одну особь.
Факторы смертности природных популяций очень разнообразны. Смертность особей может быть вызвана влиянием абиотических (низкие и высокие температуры, ливни, град, избыточная или недостаточная влажность, сильный ветер) или биотических (отсутствие корма, инфекционные заболевания, хищники, конкуренты) факторов среды. За последнее столетие лидирующую позицию среди факторов смертности заняли антропогенные факторы (загрязнение окружающей среды, разрушение местообитаний, прямое уничтожение животных и растений).

На протяжении жизненного цикла особей различают три типа смертности, для характеристики которых применяют графическое построение «кривых выживания», отражающих процент выживаемости в каждой возрастной группе одного поколения.
Кривая типа I (типичный пример — гидра) имеет почти линейную форму и характерна для организмов с равномерной смертностью на всех стадиях развития. Такой тип кривой в природе встречается крайне редко и только в оптимальных условиях среды. Однако он характерен для многих рыб, пресмыкающихся, птиц, травянистых многолетних растений, если отсчет ведется от организмов, уже прошедших наиболее уязвимые стадии своего развития.
Кривая типа II (типичный пример — устрица) имеет вогнутую форму. Она характерна для организмов, которые в основном гибнут на начальных этапах своего развития. Например, устрицы во взрослом состоянии ведут прикрепленный образ жизни, а их личинки относятся к планктону, именно в этот период они наиболее уязвимы. У особей, прошедших личиночную стадию, шанс выжить сильно повышается. Такой тип кривой выживания характерен для многих животных с высокой плодовитостью и отсутствием заботы о потомстве. Повышенная смертность в раннем возрасте характерна для большинства видов растений и пойкилотермных животных (у деревьев до зрелости доживает менее 1 % всходов, у рыб — 1—2 % мальков, у насекомых — менее 0,5 % личинок).
Кривая типа III (типичный пример — дрозофила) имеет выпуклую форму. Она описывает ситуацию, когда высокая смертность наблюдается в зрелом возрасте. Это характерно для дрозофил, поденок и других насекомых, которые вскоре после выхода из куколки оставляют потомство и погибают. К кривой типа III приближаются кривые выживания крупных млекопитающих и человека. Высокая выживаемость до старости обычно наблюдается у животных, для которых характерна забота о потомстве или чьи личиночные стадии проходят в благоприятных, мало изменяющихся условиях среды: в почве, древесине, живых организмах.
Откладывая по оси абсцисс продолжительность жизни в процентах от общей продолжительности жизни, можно сравнивать кривые выживания организмов, продолжительность жизни которых имеет значительные различия. На основании таких кривых можно определить периоды жизненного цикла, в течение которых тот или иной вид особенно уязвим. Эта информация имеет большое практическое значение в сельском хозяйстве и в природоохранной деятельности, так как позволяет правильно планировать сроки проведения мероприятий по регулированию численности популяций.
Поскольку смертность подвержена более резким колебаниям и больше зависит от факторов окружающей среды, чем рождаемость, то она играет главную роль в регулировании численности популяций. Увеличение смертности, как правило, приводит к снижению численности популяции, если нет увеличения рождаемости, и свидетельствует о неблагоприятном воздействии факторов окружающей среды.
Разница между рождаемостью и смертностью определяет рост численности популяций и позволяет прогнозировать изменение численности популяций в будущем. Количественной характеристикой роста численности является показатель скорости роста численности — изменение числа особей в популяции за единицу времени.
В случае равенства рождаемости и смертности численность популяции поддерживается на постоянном уровне — рост численности приближается к нулю, и популяция называется стабильной. Ее плотность незначительно отличается от средней величины, а ареал при этом не увеличивается и не уменьшается.
Если рождаемость превышает смертность, то численность популяции возрастает — рост численности положительный, и популяция называется растущей. Для растущих популяций характерны вспышки массового размножения (саранча, жук колорадский, грызуны, вороны, воробьи, борщевик, одуванчик). При переуплотнении у растений начинается самоизреживание, у животных — миграция на сопредельные свободные участки.
Если смертность превышает рождаемость, то численность популяции снижается — рост численности отрицательный, и такая популяция считается сокращающейся. В естественной среде она сокращается до определенного предела, а затем рождаемость, как правило, вновь повышается, и популяция из сокращающейся может стать растущей. Если этого не происходит, то популяции грозит вымирание. Чаще всего сокращающимися популяциями являются популяции редких, реликтовых, ценных как в экономическом, так и в эстетическом отношении видов.
Повторим главное. К динамическим свойствам популяции относятся рождаемость, смертность, рост численности. В оптимальных условиях среды в популяциях будет наблюдаться максимальная рождаемость и минимальная смертность. Для рождаемости и смертности можно рассчитать показатели абсолютной и удельной рождаемости и смертности. Чтобы сравнивать по рождаемости или смертности разные по численности популяции, надо использовать только показатели удельной рождаемости и смертности. Для характеристики смертности на протяжении жизни поколения используют три типа кривых выживания. Они позволяют определить периоды, в течение которых наиболее высокая смертность, а значит, тот или иной вид особенно уязвим. Разница между рождаемостью и смертностью характеризует рост численности популяции. В зависимости от характера роста численности популяции могут быть стабильными, растущими или сокращающимися.
Смертность характеризует гибель особей в популяции и выражается числом особей, погибших в единицу времени. Удельная смертность характеризует уровень смертности для определенной группы особей, например, количество смертей за месяц на тысячу особей. Смертность также зависит от факторов среды и бывает обычно значительно выше минимальной смертности при идеальных условиях среды, которая определяется физиологией данного вида организмов — даже в идеальных условиях особи будут умирать от старости.[ …]
Общая смертность — это число особей, погибающих в единицу времени (ДУУт); удельная смертность т = ДА / Д/уУ. Минимальная смертность возможна только в идеальных условиях как результат физиологической старости. Сопротивление среды повышает ее до реальной (экологической) смертности.[ …]
Абсолютная и удельная смертность характеризуют скорость убывания численности популяции вследствие гибели особей от хищников, болезней, старости и т. д.[ …]
Рождаемость и смертность, или скорость рождаемости и смертности, выражают соответственно отношениями: ANn/ At и ANm/At. Но для сравнения рождаемости или смертности в различных популяциях пользуются величиной удельной рождаемости или удельной смертности — отношением скорости рождаемости или смертности к исходной численности (N): AN/NAt и ANm/NAt. Если, например, условно принять, что в популяции инфузорий было 200 особей и через один час стало 400 при отсутствии смертности, то рост популяции за счет рождаемости составил 200 особей в час, и эту же величину можно принять за скорость рождаемости. Удельная рождаемость, т.е. в пересчете прироста на одну особь, за это же время составит 200:200×1 = 1 особь в час.[ …]
Рождаемость и смертность, или скорость рождаемости и смертности, выражают соответственно отношениями: ANn/ At и ANm/At. Но для сравнения рождаемости или смертности в различных популяциях пользуются величиной удельной рождаемости или удельной смертности — отношением скорости рождаемости или смертности к исходной численности (N): AN/NAt и ANm/NAt. Если, например, условно принять, что в популяции инфузорий было 200 особей и через один час стало 400 при отсутствии смертности, то рост популяции за счет рождаемости составил 200 особей в час, и эту же величину можно принять за скорость рождаемости. Удельная рождаемость, т.е. в пересчете прироста на одну особь, за это же время составит 200:200×1 = 1 особь в час.[ …]
Внутривидовая конкуренция ведет к зависимому от плотности увеличению смертности или к зависимому от плотности снижению рождаемости. Нарис. 7 иллюстрируется, как с увеличением плотности удельная рождаемость снижается, а удельная смертность возрастает.[ …]
За бесконечно малый промежуток времени (At —» 0) мы получим, соответственно, мгновенную удельную рождаемость, которую обозначают латинской буквой «Ъ», и мгновенную удельную смертность, которую обозначают буквой «d». Эти величины имеют размерность «единица времени-1» и зависят соответственно от интенсивности размножения или гибели особей. Например, интенсивность размножения для бактерий — час, для фитопланктона — сутки, для насекомых — неделя или месяц, для крупных млекопитающих — год.[ …]
Традиционно применяется несколько основных подходов к оценке потенциальных ОПО и связанных с ними воздействий: “концепция удельной смертности”; статистический метод (в этом случае дается оценка опасности “снизу”); оценка максимального ущерба (наиболее распространенный в практике страхования метод; дается оценка опасности “сверху”); количественная оценка риска (в основе построение множества сценариев развития возможных аварий на объекте с последующей оценкой частоты реализации каждого из сценариев и определением масштабов последствий аварий).[ …]
Читатель, возможно, уже заметил, что масштаб, отложенный на оси у на рис. 4.7,5 — логарифмический и что использование логарифмов при построении кривых выживания весьма существенно. Лучше всего пояснить это на примере двух воображаемых работ по изучению одной и той же популяции. В первом исследовании пересчитывается вся популяция, а во втором — только выборки из нее, эквивалентные одной десятой ее части (при этом о величине этой части исследователь ничего не знает). Допустим, что в первом случае результат подсчета за определенный временной интервал снижается с 1000 до 500 особей. Тогда во втором случае «индекс плотности» должен понизиться со 100 до 50. Наклон арифметической кривой выживания будет равен —500 в первом случае, но лишь —50 во втором, хотя оба они биологически идентичны, т. е. вероятность гибели одного индивидуума (удельная смертность) в обоих случаях одна и та же. Если же, однако, построить логарифмические кривые выживания, то эта идентичность будет отражена: наклон в обоих случаях составит —0,301. По этим причинам при графическом изображении изменений численности популяций мы будем часто отдавать логарифму численности предпочтение перед численностью как таковой.[ …]